Sleep is of interest to psychologists for a variety of reasons. There is the persisting puzzle of its function. Does it serve physiological restoration, mental restoration, both, or neither? There is the role of sleep in memory consolidation. For clinical psychologists, there is insomnia and other disorders of sleep, to be diagnosed and treated. I will have little to say about these matters in these lectures, which will focus on sleep as an altered state of consciousness.
Fortunately, UC Berkeley offers
other courses that deal with these
and other aspects of sleep.
However,
because most college students
-- and, for that matter, probably most college faculty
-- don't get enough sleep, I provide a couple of
questionnaires that will help you evaluate your own
sleeping behavior.
Sleepiness and Sleep ExperiencesLink to surveys of sleepiness, "sleep debt", and sleep experiences.
|
And now, on to sleep as an altered state of consciousness.
In behavioral terms, sleep appears to be a homogeneous state, one of many circadian rhythms (from the Latin circa diem, meaning "about a day") evidenced in cyclical changes in activity levels, hormonal levels, and body temperature that alternate over a period of approximately one day. Although the function of sleep is unknown, one effect of the sleep-wake cycle is to synchronize behavior and physiological states to the changing state of the environment. Thus, humans and some nonhuman animals are diurnal, meaning that they are active during the day, while other nonhuman animals are nocturnal, meaning that they are active during the night.
The sleep-wake cycle is a circadian rhythm regulated (we
think) by the suprachiasmatic nucleus (SCN) of the
hypothalamus, a neural structure that is located above the
optic chiasm (Stephan & Zucker, 1972; Moore & Eichler,
1972). The SCN receives information about the presence
of environmental light through the retinothalamic pathway,
which permits behavior and physiology to be entrained to the
actual pattern of light and dark in the environment.
Thus, environmental light serves as a Zeitgeber (from
the German, "time-giver"). People who are blind because
of damage to the eyes suffer from abnormalities in the
sleep-wake cycle, and various sleep disorders, in a way that
people who are blind from damage to the visual cortex do not.
In addition to the SCN, there may be other circadian oscillators. For example, in the absence of light cues, our internal biological clock produces its own "free-running" circadian rhythm of approximately 25 hours in length. Apparently, the Zeitgeber is reset from signals from specialized cells in the retina of the eye (cells that are different from the rods and cones involved in visual sensation).
The pineal gland, made famous by Descartes as the hypothetical link between mind and body, appears to play a role in the sleep-wake cycle, releasing melatonin ("the darkness hormone") during periods of darkness.
Circadian rhythms are a subset of a larger class of biological rhythms.The diurnal cycle of waking and sleeping seems
to be governed by two somewhat different mechanisms:
For more about the physiology of sleep,
various sleep disorders, and proper sleep hygiene, see:
The Culture of SleepSleep isn't just for physiologists and
psychologists. There's also a sociology and
anthropology of sleep. Consider, for example, what
life was like before electrification. The sun went
down candles were expensive, and so there was little to do
at night except sleep. (Well, yes, there was that,
too.) Some of the sociocultural aspects of sleep are
explored by Benjamin Reiss in Wild Nights: How Taming
Sleep Created Our Restless World (2017; reviewed by
Jennifer Senor in the New York times,
03/02/2017). Reiss has produced a social history and
anthropology of sleep -- not by looking at the scientific
and medical literature on sleep, but by looking a
literature -- including old medical textbooks.
He argues persuasively that "Virtually nothing
about our standard model of sleep existed as we know it
two centuries ago". There used to be "sleep
diversity"; now, with industrialization and
standardization, never mind the 24/7 information economy,
everybody sleeps the same way. And unequally: Reiss
has a discussion of what he refers to as "sleep
inequality" between upper and lower classes. And he
also suggests that our desire to "tame sleep" is part and
parcel of our desire to tame nature in general -- and
probably to similar effect. |
How do we recognize that sleep has occurred in others, and in ourselves?
First, sleep is manifested in our overt, gross behavior: There is the appearance of physical relaxation (e.g., the person is often lying prone), and a pattern of slow, even breathing. In fact, before the advent of sophisticated electrophysiological techniques (such as the EEG) for monitoring sleep, sleep was often diagnosed solely in terms of patterns of respiration.
Second, sleep is manifested in subjective experience. We experience, at least retrospectively, an interruption of our normal waking consciousness or awareness. There also may be a momentary disorientation upon awakening. We usually cannot remember events that transpired while we were (apparently) asleep, and we often remember having dreamed.
To the sleeper, and to the external observer, sleep appears to be a homogeneous state. But when we go beyond superficial introspection and behavioral observation, psychophysiological studies of sleep reveal that there are many different kinds of physiological changes and mental activities during sleep, and that we shift back and forth from one state to another, throughout the night, on a 90-minute cycle (i.e., an ultradian rhythm) of shifting psychophysiological states.
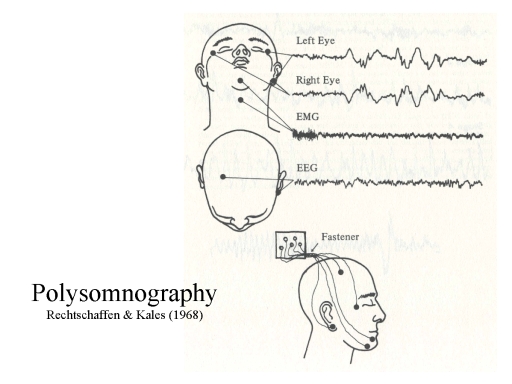 The stages of sleep are defined principally
by three physiological indices:
The stages of sleep are defined principally
by three physiological indices:
 What's
depicted above is the minimal setup for polysomnography,
involving only a single EEG channel. Nowadays, when EEG records
are displayed on a computer screen rather than paper, and
computers have a tremendous amount of processing and
data-storage capacity, most sleep laboratories record multiple
channels of EEG from electrodes placed at various locations
around the scalp, in the hopes of more discretely localizing the
source of the signals being recorded.
What's
depicted above is the minimal setup for polysomnography,
involving only a single EEG channel. Nowadays, when EEG records
are displayed on a computer screen rather than paper, and
computers have a tremendous amount of processing and
data-storage capacity, most sleep laboratories record multiple
channels of EEG from electrodes placed at various locations
around the scalp, in the hopes of more discretely localizing the
source of the signals being recorded.
 The aggregate EEG can be broken up into
various bands, waves, or rhythms, based
on their frequency (cycles per second), amplitude (microvolts),
shape, and the scalp location where they are most
prominent.
The aggregate EEG can be broken up into
various bands, waves, or rhythms, based
on their frequency (cycles per second), amplitude (microvolts),
shape, and the scalp location where they are most
prominent.
 There are lots of these EEG
rhythms, but only three are most important in the context of
sleep.
There are lots of these EEG
rhythms, but only three are most important in the context of
sleep.
|
Alpha activity: Rhythmic waves, moderate frequency 8-12 cycles per second (Hz), high amplitude 5-100 microvolts) Beta activity: Desynchronized waves, fast (18-30 Hz), low amplitude (2-20 microvolts) Delta activity: Rhythmic waves, very low frequency (0.5-4 Hz), high amplitude (20-200 microvolt) |
 The normal
waking state is characterized by a mix of alpha
and beta activity in the EEG. The relative
amounts of alpha and beta activity depend on the mental
activity of the subject. "Active looking" tends to
block (diminish) alpha activity. In addition, the
amount of alpha activity is related to arousal by an
inverted-U-shaped function: alpha is low at both low and
high levels of arousal, high at moderate levels.
The normal
waking state is characterized by a mix of alpha
and beta activity in the EEG. The relative
amounts of alpha and beta activity depend on the mental
activity of the subject. "Active looking" tends to
block (diminish) alpha activity. In addition, the
amount of alpha activity is related to arousal by an
inverted-U-shaped function: alpha is low at both low and
high levels of arousal, high at moderate levels.
 Drowsiness is
characterized by an increase (as the eyes close or defocus),
and then a decrease in EEG alpha. As the sleeper moves
down a continuum of arousal, the EMG shows signs of decreasing
activity in the skeletal nervous system (SNS), including
relaxation of muscle tension and decreasing bodily
motility. There are also signs of relaxation in the
autonomic nervous system (ANS), including reduced heart rate,
respiration, and body temperature.
Drowsiness is
characterized by an increase (as the eyes close or defocus),
and then a decrease in EEG alpha. As the sleeper moves
down a continuum of arousal, the EMG shows signs of decreasing
activity in the skeletal nervous system (SNS), including
relaxation of muscle tension and decreasing bodily
motility. There are also signs of relaxation in the
autonomic nervous system (ANS), including reduced heart rate,
respiration, and body temperature.
The sleeper's eyes eventually close, but the onset of sleep is actually defined by the disappearance of alpha activity from the EEG.
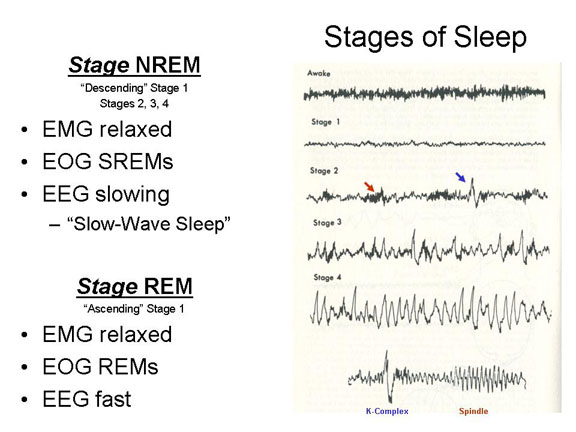 Descending Stage
1 sleep is defined by an alpha-free EEG, dominated by
low-voltage, fast, desynchronized beta-like activity
resembling the waking EEG, except for a somewhat lower
amplitude (voltage) -- partly, perhaps, because of the absence
of high-voltage alpha activity in the mix. In this and
the remaining stages of sleep, through Stage 4, the EMG signs
of increasing relaxation in the SNS. The EOG also shows
slowly oscillating (rolling) eye movements (SREMs) through
this entire period.
Descending Stage
1 sleep is defined by an alpha-free EEG, dominated by
low-voltage, fast, desynchronized beta-like activity
resembling the waking EEG, except for a somewhat lower
amplitude (voltage) -- partly, perhaps, because of the absence
of high-voltage alpha activity in the mix. In this and
the remaining stages of sleep, through Stage 4, the EMG signs
of increasing relaxation in the SNS. The EOG also shows
slowly oscillating (rolling) eye movements (SREMs) through
this entire period.
In a behavioral test, the subject's lack of responsiveness confirms that the person is in fact asleep. However, upon awakening from this stage people will frequently deny having been asleep.
Stage 1 is a transitional phase in the sleep cycle, usually accounting for about 5% of a night's sleep.
 Stage 2, the
next stage "down" in sleep, is characterized by a continued
deepening of relaxation and SREMs. The EEG is
characterized by spindles (bursts of 14-18 Hz activity) and
K-complexes (paroxysmal bursts of high-amplitude
activity). The spindles reflect a burst of electrical
activity that may help bind recently acquired information into
the pre-existing network of knowledge stored in memory.
When subjects learn a new task during the day, the amount of
spindle activity increases that night; and the amount of
spindle activity the night before is correlated with
performance on the new task the next day.
Stage 2, the
next stage "down" in sleep, is characterized by a continued
deepening of relaxation and SREMs. The EEG is
characterized by spindles (bursts of 14-18 Hz activity) and
K-complexes (paroxysmal bursts of high-amplitude
activity). The spindles reflect a burst of electrical
activity that may help bind recently acquired information into
the pre-existing network of knowledge stored in memory.
When subjects learn a new task during the day, the amount of
spindle activity increases that night; and the amount of
spindle activity the night before is correlated with
performance on the new task the next day.
Stage 2, with the disappearance of alpha activity in the EEG, and the appearance of spindles and K-complexes, marks the true onset of sleep, and consumes about 45% of the sleeper's time during the night.
 Stage 3 is
characterized by more profound relaxation in the EMG, and the
continuation of SREMs. The EEG shows the first
appearance of delta activity. Awakening the subject
requires a loud noise, or perhaps repetition of the subject's
name.
Stage 3 is
characterized by more profound relaxation in the EMG, and the
continuation of SREMs. The EEG shows the first
appearance of delta activity. Awakening the subject
requires a loud noise, or perhaps repetition of the subject's
name.
 Stage 4 is
dominated by EEG delta activity (more than half the
time). The person's muscles are very relaxed, and he or
she rarely moves. It is very difficult to awaken the
subject (the technical term for this is "high auditory arousal
threshold"). If awakened, the subject feels very groggy,
and comes into focus only very slowly -- a phenomenon known as
sleep inertia.
Stage 4 is
dominated by EEG delta activity (more than half the
time). The person's muscles are very relaxed, and he or
she rarely moves. It is very difficult to awaken the
subject (the technical term for this is "high auditory arousal
threshold"). If awakened, the subject feels very groggy,
and comes into focus only very slowly -- a phenomenon known as
sleep inertia.
 Stage REM occurs after sleep moves
"upward", back through stages 3 and 2, and ascends into a new
stage, which resembles Stage 1 in some ways.
Desynchronized beta activity dominates the EEG, and alpha
activity is absent (otherwise the person would be
awake). The EOG shows bursts of rapid
conjugate eye movements (REMs), as if the sleeper were looking
at something (though the eyes remain closed), which gives this
stage its name.
Stage REM occurs after sleep moves
"upward", back through stages 3 and 2, and ascends into a new
stage, which resembles Stage 1 in some ways.
Desynchronized beta activity dominates the EEG, and alpha
activity is absent (otherwise the person would be
awake). The EOG shows bursts of rapid
conjugate eye movements (REMs), as if the sleeper were looking
at something (though the eyes remain closed), which gives this
stage its name.
There are also signs of autonomic arousal, including bursts of high heart rate and irregular respiration. In males, there is commonly erection of the penis. In females there is an increase in vaginal blood flow.
And, of course, subjects who are awakened during REM are very likely to report that they were dreaming.
 In Stage REM, however, the EMG still shows
signs of deep muscle relaxation, virtually a paralysis, known
as motor atonia -- at least below the
neck. There is a central inhibition of those parts of
the skeletal musculature mediated by the spinal cord (i.e.,
the spinal nerves), though not necessarily those muscles
served by the cranial nerves (or else REMs could not
occur). This paralysis effectively prevents sleepers
from acting out their dreams. But it can sometimes
continue after awakening, resulting in the relatively rare
phenomenon of sleep paralysis, which terminates either
spontaneously or after touching the subject or calling his or
her name. (Sleep paralysis is commonly thought to be
implicated in many reports of alien abduction.)
In Stage REM, however, the EMG still shows
signs of deep muscle relaxation, virtually a paralysis, known
as motor atonia -- at least below the
neck. There is a central inhibition of those parts of
the skeletal musculature mediated by the spinal cord (i.e.,
the spinal nerves), though not necessarily those muscles
served by the cranial nerves (or else REMs could not
occur). This paralysis effectively prevents sleepers
from acting out their dreams. But it can sometimes
continue after awakening, resulting in the relatively rare
phenomenon of sleep paralysis, which terminates either
spontaneously or after touching the subject or calling his or
her name. (Sleep paralysis is commonly thought to be
implicated in many reports of alien abduction.)
![]() Stages
2-4 are collectively known as Stage NREM, owing to the
absence (N) of rapid eye movements. It is also known as
slow-wave sleep (SWS), because all three stages
are characterized by relatively low-frequency activity in the
EEG. The intervening stage, of course, is known as Stage
REM. Before the discovery of REMs, the combination
of EEG and autonomic arousal characteristic of waking with EMG
characteristic of sleeping led investigators to call
"ascending" Stage 1 paradoxical sleep. It seems
likely that SWS serves the hypothesized restorative function
of sleep: it's when the body and brain appear to be most
inactive. This is especially the case for Stage 4, which
in electrophysiological terms resembles a coma -- a coma into
which we spontaneously enter, and out of which we
spontaneously emerge, several times per night.
Stages
2-4 are collectively known as Stage NREM, owing to the
absence (N) of rapid eye movements. It is also known as
slow-wave sleep (SWS), because all three stages
are characterized by relatively low-frequency activity in the
EEG. The intervening stage, of course, is known as Stage
REM. Before the discovery of REMs, the combination
of EEG and autonomic arousal characteristic of waking with EMG
characteristic of sleeping led investigators to call
"ascending" Stage 1 paradoxical sleep. It seems
likely that SWS serves the hypothesized restorative function
of sleep: it's when the body and brain appear to be most
inactive. This is especially the case for Stage 4, which
in electrophysiological terms resembles a coma -- a coma into
which we spontaneously enter, and out of which we
spontaneously emerge, several times per night.
Although muscle relaxation is a prominent feature of REM
sleep, it occurs in NREM sleep as well. In a study of
mice, Liu et al. (Science, 01/24/2020) noted activity
in the substantia nigra pars reticulata (SNr) both induced
NREM (effectively, Stage 1) sleep and inhibited movemen via
connections to the motor thalamus. This pathway is quite
different from the one that induces motor atonia in REM sleep,
which involves brain-stem circuits that suppress activity in
the spinal cord.
 The
sleep cycle consists of a succession of these stages, each
epoch consisting of Stage NREM followed by Stage REM.
The
sleep cycle consists of a succession of these stages, each
epoch consisting of Stage NREM followed by Stage REM.
 Each epoch takes approximately
90 minutes. Thus, in eight hours of sleep, a person
would go through approximately 5 complete epochs, typically
awakening out of Stage REM.
Each epoch takes approximately
90 minutes. Thus, in eight hours of sleep, a person
would go through approximately 5 complete epochs, typically
awakening out of Stage REM.There are also interesting developmental trends in the
sleep cycle.
 As
indicated in the lectures on Anesthesia and
Coma, polysomnography should greatly improve the
differential diagnosis of coma and the persistent vegetative
state. Comatose patients should not show any sleep cycles,
whether behaviorally (the usual criterion for clinical
diagnosis) or physiologically (which is what polysomnography
does). See Cologan et al. (2009) for a review.
As
indicated in the lectures on Anesthesia and
Coma, polysomnography should greatly improve the
differential diagnosis of coma and the persistent vegetative
state. Comatose patients should not show any sleep cycles,
whether behaviorally (the usual criterion for clinical
diagnosis) or physiologically (which is what polysomnography
does). See Cologan et al. (2009) for a review.
 We'll have more to say about the
physiology of the 90-minute cycle of waking and sleeping in the
Lecture Supplement on "Dreaming".
To make a long story short: REM is regulated by PGO waves,
bursts of neural activity that arise in the pons, and then
pass through the lateral geniculate nucleus of the
thalamus. From there they end up in the visual areas of the
occipital lobe, and produce the vivid visual images that are
characteristic of dreams. At the same time, there is activity
in the motor cortex, which is prevented by inhibitory centers in the
pons from reaching the spinal cord and the skeletal musculature.
We'll have more to say about the
physiology of the 90-minute cycle of waking and sleeping in the
Lecture Supplement on "Dreaming".
To make a long story short: REM is regulated by PGO waves,
bursts of neural activity that arise in the pons, and then
pass through the lateral geniculate nucleus of the
thalamus. From there they end up in the visual areas of the
occipital lobe, and produce the vivid visual images that are
characteristic of dreams. At the same time, there is activity
in the motor cortex, which is prevented by inhibitory centers in the
pons from reaching the spinal cord and the skeletal musculature.
So, sleep is something we do, and we do it every day,
pretty much like clockwork. Every organism with a
nervous system has a sleep-wake cycle (and even many flowers
close up for the night!). Maybe sleep is just a
holdover from our phylogenetic heritage. But why
do we do it? Surprisingly, we don't really have very
many good ideas about this.
Every animal sleeps. Even jellyfish, who don't even
have a brain, appear to sleep (if you define sleep as a
state of diminished mobility and responsiveness from which
the organism can be easily aroused). Dolphins sleep,
turning off one hemisphere at a time so they can keep
swimming; so, perhaps, do frigatebirds, so they can keep
flying.
First, it's clear that we have something like a biological need for sleep. Following a period of sleep-deprivation, people perform poorly on various mental and physical tasks, they're less alert, and they have more negative moods. And when they're allowed to sleep, they fall asleep quickly. If they're deprived of REM or NREM sleep, they'll spend more time the next night in REM -- a phenomenon known as REM rebound. There's a similar rebound for NREM sleep.
A series of experiments by David Dinges and his colleagues at the University of Pennsylvania dramatically reveals the effects of sleep-deprivation on the performance of cognitive tasks. In one study, the longest sleep-restriction study conducted to date, they had subjects spend two weeks in the laboratory, and allowed different groups a full 8 hours' sleep, or only 6 or 4 hours. Every two hours (except when they were sleeping), the subjects completed a 10-minute psychomotor vigilance task (PVT), commonly used in studies of sustained attention, in which subjects must respond whenever a number appears on a computer screen. Even short delays in responding, on the order of 500 milliseconds, indicate microsleep -- that is, that the subject has fallen asleep, however briefly. Subjects who got a "full" eight hours of sleep performed consistently well on this task, responding within a half-second to the stimuli. But the subjects who only got four or six hours performed significantly worse, even on the first day, and their performance deteriorated progressively as the two-week trial went on. By the end of the two-week session, subjects who got only six hours of sleep per might were performing as poorly as those who had been sleep-deprived for 24 hours straight. Similar research by George Belenky found that subjects who got 9 hours of sleep per night did no better on the PVT than those who got 8 hours of sleep; but those who got only 7 hours showed the same cumulative performance decrement observed in Dinges' 6-hour subjects.
A survey by the National Sleep Foundation indicates that the average American gets only about 6.9 hours of sleep per night, indicating that all too many of us are walking around sleep-deprived. Adolescents -- this means many of you -- need more sleep time. The elderly appear to get along with less sleep than the young or middle-aged -- though those who sleep longer tend to stay healthier, both physically and mentally.
But what exactly do we get from sleep? The obvious answer is that sleep performs some sort of restorative function, allowing us to recover from the day's physical activity
All of these proposals are eminently reasonable, and quite likely at least one of them is true. But nailing down the exact biological function of sleep hasn't been easy.
Turning the Tables on AdaptationismWith the possible exception of altruism, nothing bedevils evolutionary psychology more than sleep (and dreams). Why should we do something that renders us vulnerable to predation for roughly 1/3 of every day? Why should we have dreams, when we can't remember them?
We usually think of sleep as an altered state of consciousness, and philosophers often point to "dreamless sleep" as a contrast to normal, waking, consciousness. But maybe not: Maybe sleep is the normal state. Commenting on a series of articles on "The Secrets of Sleep", appearing in the National Geographic magazine, Walter Weller wrote:
|
In addition to these biological functions, sleep may also have cognitive functions. In particular, there has also been considerable interest in the effect of sleep on memory for information learned while the subject is awake.
 Consider, first, an early study
conducted by Jenkins & Dallenbach (1924) as a test of
the consolidation theory of forgetting. Subjects
learned a list of nonsense syllables, and then were tested
after retention intervals of 0-8 hours. Some of the
subjects remained awake during this period of time, others
went to sleep. Memory was actually better when
subjects slept between study and test, compared to trials
in which they stayed awake for an equivalent
interval. Based on these results, Jenkins and
Dallenbach proposed that forgetting is caused by
interference, rather than the mere passage of time.
Because "nothing" happens during sleep, less interference
occurs.
Consider, first, an early study
conducted by Jenkins & Dallenbach (1924) as a test of
the consolidation theory of forgetting. Subjects
learned a list of nonsense syllables, and then were tested
after retention intervals of 0-8 hours. Some of the
subjects remained awake during this period of time, others
went to sleep. Memory was actually better when
subjects slept between study and test, compared to trials
in which they stayed awake for an equivalent
interval. Based on these results, Jenkins and
Dallenbach proposed that forgetting is caused by
interference, rather than the mere passage of time.
Because "nothing" happens during sleep, less interference
occurs.
But in addition to preventing interference, sleep may actively enhance memory by promoting the consolidation process.
 In fact, neurophysiological studies of
rats have examined the role that sleep may play in the
consolidation of memory. McNaughton and his
colleagues (Wilson & McNaughton, 1994; Skaggs &
McNaughton, 1995) made single-unit recordings of neural
activity while rats were exposed to a spatial-learning
task. The rats were placed in a pool of water, and
had to swim around until they located a platform on which
they could rest. Over time, the animals learn to
find the platform fairly quickly. Using this
procedure, the researchers identified an ensemble of
neurons in a particular area of the hippocampus (CA1)
whose activity changed during learning -- an area
representing the spatial location of the platform.
They then examined the activity of these neurons before
and during learning, and during a post-learning period of
sleep. The same neurons were active during
"slow-wave" sleep (essentially, Stage NREM) as were active
during learning -- suggesting that consolidation is
mediated, in part, by the reactivation of memories during
sleep. As Wilson & McNaughton (1994) put it:
In fact, neurophysiological studies of
rats have examined the role that sleep may play in the
consolidation of memory. McNaughton and his
colleagues (Wilson & McNaughton, 1994; Skaggs &
McNaughton, 1995) made single-unit recordings of neural
activity while rats were exposed to a spatial-learning
task. The rats were placed in a pool of water, and
had to swim around until they located a platform on which
they could rest. Over time, the animals learn to
find the platform fairly quickly. Using this
procedure, the researchers identified an ensemble of
neurons in a particular area of the hippocampus (CA1)
whose activity changed during learning -- an area
representing the spatial location of the platform.
They then examined the activity of these neurons before
and during learning, and during a post-learning period of
sleep. The same neurons were active during
"slow-wave" sleep (essentially, Stage NREM) as were active
during learning -- suggesting that consolidation is
mediated, in part, by the reactivation of memories during
sleep. As Wilson & McNaughton (1994) put it:
"[D]uring slow-wave] sleep, the activity of hippocampal cells engaged in place-specific activity during prior waking retains the structure of distributed representations of the visited locations.... [I]nitial storage of event memory occurs through rapid synaptic modification, primarily within the hippocampus.... During… slow-wave sleep, synaptic modification within the hippocampus itself is suppressed and the neuronal states encoded within the hippocampus are "played back" as part of a consolidation process by which hippocampal information is gradually transferred to the neocortex".
Interestingly, however, the kind of sleep that seems to be involved is "slow-wave" sleep, analogous to Stage NREM. Assuming that rats have rat-dreams during REM, it's possible that the relative mental quiescence of NREM sleep is critical for consolidation.
Recently, studies of humans have strengthened the case for memory consolidation during sleep. Stickgold (2005) has reviewed a number of studies involving procedural learning -- discriminating foreground from background, motor sequencing, and motor adaptation -- in which subjects showed further improvements in performance, even after formal training had ceased, after subjects had one night of sleep, compared to a control period in which they stayed awake. (Much of this research was performed in collaboration with UCB's Prof. Matt Walker, who has now extended the essential findings to declarative learning as well.)
One experiment involved a visual-texture
discrimination task, in which subjects had to identify the
orientation of an array of diagonal bars presented against a
horizontal background. (a) When retested 6, 12, 18, or 24 hours after initial training, subjects showed further improvement if the post-training interval contained a period of sleep. |
 |
Another involved motor-sequence learning, in
which subjects were instructed to type a sequence of 1-digit
numbers on a computer keyboard.(d) Again, there was more post-training improvement in performance following an interval of sleep. |
 |
And yet a third task involved motor
adaptation, in which subjects traced a line between two
points on a computer screen against a.countervailing virtual
"force".(g) As in the other two experiments, post-training performance improved further after a night's sleep. |
 |
Interestingly, SWS (Stage NREM) appears to play a large role in these consolidation effects, compared to REM. This is congruent with McNaughton's rat study.
The function of sleep poses a big problem for
biological psychologists, but in this course we are
primarily interested in the effects of sleep on mental
life. Sleep (at least SWS) is often referred to as
a state of unconsciousness, but anecdotal reports
suggest that some kind of mental activity persists, even
when we are asleep.
Note: there is a prominent theory that incubation as such -- that is, unconscious problem-solving -- does not occur; rather, on the second trial subjects simply start the problem-solving process from a different "entry point". But that can't explain these results. All of the unsolved problems would have received the benefit of a new entry point on the second day, but still the ones whose memories were reactivated were solved more successful. So incubation does appear to occur after all, and it's facilitated by "sleeping on" unsolved problems -- provided that memories of those problems are reactivated during the night.
Falling Asleep
and Waking Up
While the function of sleep remains a mystery, what interests us in this course is sleep as an altered state of consciousness -- a radical departure from the wakeful awareness that we experience in the ordinary course of everyday living. Which brings us to the question: what is consciousness -- particularly cognition like during sleep? Is there any at all?
 Webb (1975) has noted that subjects
are more likely to report that they were asleep
if awakened from Stages 2, 3, and 4 than from
descending Stage 1 -- even though they are
actually asleep, as indicated by the absence of
alpha activity in the EEG.
Webb (1975) has noted that subjects
are more likely to report that they were asleep
if awakened from Stages 2, 3, and 4 than from
descending Stage 1 -- even though they are
actually asleep, as indicated by the absence of
alpha activity in the EEG.
 The cognitive changes
occurring while falling asleep are illustrated by a study
performed by Wyatt et al. (1994). After lights out, subjects
were presented with 16 pairs of word associates (e.g.,
BREAD-BUTTER) at the rate of 1 per minute while they were
falling asleep, and instructed to repeat each word pair
aloud after hearing it. After the subjects no longer
responded, and alpha was absent from their EEG for at least
15 seconds, presentation was discontinued and the subject
was allowed to sleep for another 30 sec or 10 min. At this
point, the subject was awakened, and asked to recall and
recognize the word pairs s/he had heard. Control subjects
were instructed to stay awake during list
presentation. This procedure was repeated for 10
cycles -- which must have been extremely annoying to those
subjects who were trying to go to sleep!
The cognitive changes
occurring while falling asleep are illustrated by a study
performed by Wyatt et al. (1994). After lights out, subjects
were presented with 16 pairs of word associates (e.g.,
BREAD-BUTTER) at the rate of 1 per minute while they were
falling asleep, and instructed to repeat each word pair
aloud after hearing it. After the subjects no longer
responded, and alpha was absent from their EEG for at least
15 seconds, presentation was discontinued and the subject
was allowed to sleep for another 30 sec or 10 min. At this
point, the subject was awakened, and asked to recall and
recognize the word pairs s/he had heard. Control subjects
were instructed to stay awake during list
presentation. This procedure was repeated for 10
cycles -- which must have been extremely annoying to those
subjects who were trying to go to sleep!
 Tests of recall
and recognition indicated a sharp drop-off in memory for the
items presented during the final three minutes of sleep
onset -- including items that the subjects had repeated
while they were still awake. Memory was especially
poor for subjects who were allowed to sleep for 10 minutes
after presentation of the last item. Recognition was
somewhat better than recall, of course, but there was still
a big difference between sleepers and controls, and
especially after 10 minutes of sleep. Wyatt et al.
suggested that these findings indicated a temporally graded
retrograde amnesia induced by sleep onset.
Tests of recall
and recognition indicated a sharp drop-off in memory for the
items presented during the final three minutes of sleep
onset -- including items that the subjects had repeated
while they were still awake. Memory was especially
poor for subjects who were allowed to sleep for 10 minutes
after presentation of the last item. Recognition was
somewhat better than recall, of course, but there was still
a big difference between sleepers and controls, and
especially after 10 minutes of sleep. Wyatt et al.
suggested that these findings indicated a temporally graded
retrograde amnesia induced by sleep onset.
 And of course,
the next thing to determine was whether this sleep-onset
amnesia dissociated explicit and implicit memory. For
the explicit memory test, Wyatt presented the cue term and
asked subjects to respond with the item with which it had
been paired. For the implicit memory test, subjects
were asked to respond simply with the first word that came
to mind. Wyatt found that subjects in the sleep
condition performed equally poorly on both explicit and
implicit tests of memory (control subjects showed a
significant priming effect). So, sleep-onset amnesia
impairs both explicit and implicit memory.
And of course,
the next thing to determine was whether this sleep-onset
amnesia dissociated explicit and implicit memory. For
the explicit memory test, Wyatt presented the cue term and
asked subjects to respond with the item with which it had
been paired. For the implicit memory test, subjects
were asked to respond simply with the first word that came
to mind. Wyatt found that subjects in the sleep
condition performed equally poorly on both explicit and
implicit tests of memory (control subjects showed a
significant priming effect). So, sleep-onset amnesia
impairs both explicit and implicit memory.
Ellenbogen and his colleagues have performed an analogous study of the process of waking up (Current Biology, 2010). Subjects slept for three nights in the laboratory, the first being a baseline night. The next two nights, they played various common sounds in the bedroom -- the sound of a toilet flushing, or a phone ringing, people talking, etc. -- as soon as they fell asleep, beginning at low volume and increasing until they awakened. The more baseline spindle activity the subjects showed during Stage 2, the more difficult it was to awaken them. The investigators suggested that spindle activity has the function of shutting out sounds from the external environment.
 Aserinsky and
Kleitman (1953) first observed the cyclical shift from
SWS to REM, with bursts of REM, lasting 10-20 minutes,
occurring approximately every 90 minutes. On
Aserinsky's hunch, they awakened sleeping subjects, and
asked them a simple question: "Were you
dreaming?".
Aserinsky and
Kleitman (1953) first observed the cyclical shift from
SWS to REM, with bursts of REM, lasting 10-20 minutes,
occurring approximately every 90 minutes. On
Aserinsky's hunch, they awakened sleeping subjects, and
asked them a simple question: "Were you
dreaming?".
 The result
was a much higher frequency of dream reports during
Stage REM (74% of awakenings) than Stage NREM (only 17%
awakenings). So, Aserinsky and Kleitman concluded that
dreams do not occur throughout sleep (as had been
suggested by Freud, for example), but are largely
confined to REM. When William Dement, a sleep
researcher at Stanford, reviewed this literature in
1976, a number of studies had confirmed
A&K's findings -- though, as we will see, there is
some controversy here.
The result
was a much higher frequency of dream reports during
Stage REM (74% of awakenings) than Stage NREM (only 17%
awakenings). So, Aserinsky and Kleitman concluded that
dreams do not occur throughout sleep (as had been
suggested by Freud, for example), but are largely
confined to REM. When William Dement, a sleep
researcher at Stanford, reviewed this literature in
1976, a number of studies had confirmed
A&K's findings -- though, as we will see, there is
some controversy here.
What is the connection between dreams and REMs? According to the scanning hypothesis, REMs reflect visual activity on the part of the dreamer -- the dreamer is actually scanning the visual world represented by the dream. On the other hand, even congenitally blind people show REMs, and when awakened from REM sleep report dreaming. So, to be more precise, we should say that according to the scanning hypothesis the dreamer is scanning the spatial world represented by the dream. Or, maybe, REMs don't have anything to do with looking at all -- they're just an adventitious consequence of nervous system activity during a particular stage of sleep.
Before Aserinsky and Kleitman, nobody had ever made all-night EEG recordings of sleeping individuals. The EEG changes associated with falling asleep -- passing from Descending Stage 1 through Stages 2 and 3 to Stage 4 -- were known. But nobody had carried the observation further, on the assumption that the brain quieted down as the person fell asleep, and remained quiet until the person woke up. Also, at the time computers were not widely available, so that polysomnographic recordings had to be made on huge (and expensive) rolls of graph paper. Recording further was believed by everyone -- including Kleitman himself! -- to be a simple waste of paper. But Aserinsky did it anyway, and after that first night everyone in Kleitman's lab became very interested.
In the end, Aserinsky's discovery had two implications:
The discovery of two distinct types of sleep, REM and NREM, showed that sleep was not a passive state characterized by the relative absence of neural activity. Instead, the brain remains active during sleep, cycling between two distinct states -- one of relative activity (REM), the other of relative inactivity (NREM).The discovery of the association between REMs and dreams showed that, despite appearances, the sleeper is not unconscious after all. But of course we've known about dreams for a long time -- they're described in myths and Bible stories, for example. Nobody needed Aserinsky and Kleitman to tell them that they dreamt while they were asleep. The real achievement of Aserinsky and Kleitman was to tie dreaming to a specific aspect of physiology -- REMs. By tying dreams to something other than the subject's unverifiable self-report, Aserinsky and Kleitman made the scientific study of dreams possible.
And by showing that REMs, and thus dreams, occurred periodically, reflecting an intrinsic biological rhythm, Aserinsky and Kleitman began to undermine the Freudian, psychoanalytic theory of dreams as symbolic expressions of repressed instinctual urges. Instead, they appear to be the automatic consequence of a particular cycle of biological activity, occurring largely independently of what the person thinks, feels, and desires.
Kleitman, Azerinsky, and Dement
Aserinsky provided his own account of his discovery in the Journal of the History of the Neurosciences, 1996; see also "The Stubborn Scientist Who Unraveled a Mystery of the Night", Smithsonian, 10/03. |
The early findings of Aserinsky and Kleitman were subsequently replicated by a number of other laboratories, and so the traditional conclusion has been that dreams occur in Stage REM. However, it's not completely clear that dreams are solely a phenomenon of REM. However, as we'll see in the lectures on Dreams, the situation is actually a little bit more complicated than that -- and the complications have serious implications for our understanding of the nature of dreams.
As noted, nightmares are a special category of dreams, defined by their content and their consequences: they evoke anxiety, and they awaken the dreamer. Nightmares tend to occur late in the night's sleep.
 The
term nightmare is derived from Old English: a
mare is a spirit that suffocates a sleeping
person or other animal. In medieval mythology,
there were actually two of these: the incubus,
a female demon who gathered semen from men while they
slept; and the succubus, a male demon who
deposited the semen in women while they slept.
The classic clinical study of nightmares is Ernest
Hartmann's The Nightmare (1984). And the
classic artistic depiction is Henry Fuseli's "The
Nightmare" (1781, now in the Detroit Institute
of Art), which painting depicts an incubus sitting on
a sleeping woman. What's the horse doing there?
Maybe it's a pun -- night mare, get it?
Or maybe not. For a discussion of the painting
(a copy of which graced Freud's study in Vienna),see
"Mystery Stalks a Haunting Dream" by Johm J. Miller, Wall
Street Journal, 10/31/2020.
The
term nightmare is derived from Old English: a
mare is a spirit that suffocates a sleeping
person or other animal. In medieval mythology,
there were actually two of these: the incubus,
a female demon who gathered semen from men while they
slept; and the succubus, a male demon who
deposited the semen in women while they slept.
The classic clinical study of nightmares is Ernest
Hartmann's The Nightmare (1984). And the
classic artistic depiction is Henry Fuseli's "The
Nightmare" (1781, now in the Detroit Institute
of Art), which painting depicts an incubus sitting on
a sleeping woman. What's the horse doing there?
Maybe it's a pun -- night mare, get it?
Or maybe not. For a discussion of the painting
(a copy of which graced Freud's study in Vienna),see
"Mystery Stalks a Haunting Dream" by Johm J. Miller, Wall
Street Journal, 10/31/2020.
The findings of Aserinsky and Kleitman showed clearly that the sleeper is not unconscious after all -- at least not in Stage REM. But what about Stage NREM? Possibly the sleeper is unconscious in this stage of sleep, but NREM awakenings do typically yield reports of isolated thoughts and images, and REM-awakening studies do yield some clear dream reports after NREM awakenings as well. Dement and Kleitman (1957) solved the problem by suggesting that NREM "dreams" were really just memories of dreams from a previous REM state (Dement, like Aserinsky, was a student in Kleitman's lab, and went on to found and direct the Sleep Research Laboratory at Stanford University).
If NREM "dreams" are really memories of REM dreams, then it might be the case that the sleeper is actually unconscious in Stage NREM after all. But it is now clear that the sleeper is not unconscious even in NREM. Still, the consensus view is that NREM mentation is typically non-dreamlike thinking, reverie, and static imagery.
 The
images, reveries, and daydreams characteristic of
Descending Stage 1 have a special name: the hypnagogic
state (Schacter, 1976). In this state,
the person experiences abundant, vivid visual and
auditory imagery -- less bizarre than in dreams,
somewhat fleeting and disorganized, and with flat
affect; but still, abundant and vivid imagery.
The
images, reveries, and daydreams characteristic of
Descending Stage 1 have a special name: the hypnagogic
state (Schacter, 1976). In this state,
the person experiences abundant, vivid visual and
auditory imagery -- less bizarre than in dreams,
somewhat fleeting and disorganized, and with flat
affect; but still, abundant and vivid imagery.
Although hypnagogic imagery appears virtually full-blown, and requires little time to develop, its content changes over time as the individual, falling asleep, diminishes contact with reality.
Before Sleep and DreamsThe hypnagogic state is portrayed musically in "Before Sleep and Dreams", a 1990 piece by the American composer Aaron Jay Kernis. The piece attempts to capture the state of being half-awake, with hints of an image -- particularly, Debussy's "Engulfed Cathedral". |
These "night terrors" , also known as incubus (a name inspired by a famous painting, shown earlier, of a sleeping woman with a winged devil sitting on her torso), are usually observed in children (Gastaut & Broughton, 1966). The child will awaken with an inarticulate scream, often followed by a cry for help, and an expression of fear, accompanied by signs of an intense discharge of autonomic nervous system activity (sweating, shivering, weeping). Upon awakening, the child is unable to give an account of his or her distress. Typically, the child will fall back asleep, sleep peacefully through the night, and have no memory of the incident in the morning.
 Night terrors are distinct from
nightmares, which are really just REM dreams with
themes of anxiety or catastrophe (danger, injury,
death, social affront, embarrassment). Upon
awakening from a nightmare, the sleeper can give
an account of the dream, and remember it in the
morning. While nightmares occur in Stage
REM, usually toward the end of the night's sleep,
night terrors usually occur in periods of arousal
from Stages 3 or 4 (i.e., moving toward Stage 2),
early in the night.
Night terrors are distinct from
nightmares, which are really just REM dreams with
themes of anxiety or catastrophe (danger, injury,
death, social affront, embarrassment). Upon
awakening from a nightmare, the sleeper can give
an account of the dream, and remember it in the
morning. While nightmares occur in Stage
REM, usually toward the end of the night's sleep,
night terrors usually occur in periods of arousal
from Stages 3 or 4 (i.e., moving toward Stage 2),
early in the night.
 In sleepwalking, the sleeper
engages in activity that resembles waking life
(Kales et al., 1966a, 1966b). On closer
inspection, these behaviors are typically simple,
random, and apparently purposeless -- aimless
walking, opening doors, turning lights or the
television on and off, fumbling with
objects. Occasionally the sleeper engages in
more complex episodes. Sleepwalking is
occasionally accompanied by talking, or at least
incoherent mumbling. The sleepwalker
typically does not display any affect, and has a
"dazed" appearance.
In sleepwalking, the sleeper
engages in activity that resembles waking life
(Kales et al., 1966a, 1966b). On closer
inspection, these behaviors are typically simple,
random, and apparently purposeless -- aimless
walking, opening doors, turning lights or the
television on and off, fumbling with
objects. Occasionally the sleeper engages in
more complex episodes. Sleepwalking is
occasionally accompanied by talking, or at least
incoherent mumbling. The sleepwalker
typically does not display any affect, and has a
"dazed" appearance.
Sleepwalking typically occurs in Stage NREM, usually in an early episode of Stage 4. Of course, sleepwalking during REM would be generally prohibited by the muscle paralysis that is one of the defining features of that stage. However, motor activity can occur in REM, due to incomplete muscle paralysis (see below).
An episode of sleepwalking does not always progress to actual walking. If the person physically relocates, the episode typically ends in a spontaneous awakening. If there is no physical relocation, the sleeper resumes his or her normal sleep cycle uninterrupted. In either case, the sleepwalker has no memory for the episode.
It has been suggested that sleepwalking comes in two types:
La SonnambulaSomnambulism
provides the plot for
La Sonnambula,
an opera by Vincenzo
Bellini (1831) --
pictured here is a
publicity still from
Metropolitan Opera
production, with
Natalie Dessay in the
title role, broadcast
03/21/2009.
For more background, see "Sleep Disorder" by Steven Blier, Opera News, 03/2009. |
Sleeptalking refers to the utterance of speech, and other meaningful sounds, by someone who is asleep (Arkin, 1978). If the sleeptalker is awakened, he or she will have no awareness of talking.
Sleeptalking is almost ubiquitous, though it occurs more often in children than in adults.
Sleeptalking typically occurs during periods of transient arousal from NREM sleep, though it can also occur in REM.
Roommates take note: sleeptalkers
are fun to listen to, but they rarely reveal any
secrets.
Parasomnias
Sleepwalking and night terrors are part of a larger category of sleep disorders or parasomnias, which include various forms of insomnia: initial insomnia (difficulty falling asleep), middle insomnia (awakening after falling asleep, and difficulty getting back to sleep), and terminal insomnia (awakening too early). For a literary account of insomnia, see Wide Awake: A Memoir of Insomnia by Patricia Morrisoe (2010), whose own story provides a framework for a wide-ranging discussion of both normal and pathological aspects of sleep -- and the pharmaceutical "insomnia industry". See also The Family that Couldn't Sleep by D.T. Max, which focuses on a genetic disorder known as fatal familial insomnia -- which is just what it sounds like. Speaking of which, a large number of pharmaceutical sleep aids drugs are available, either by prescription or over the counter, for the treatment of insomnia. Barbiturate and benzodiazepine drugs are not generally used for this purpose, as they inhibit REM sleep. Instead, physicians will often prescribe nonbenzodiazepine hypnotics such as Lunesta, Sonata, and Ambien. These sleep aids appear to work primarily by enhancing the activity of a neurotransmitter known as GABA. In addition to sleepwalking and night terrors, parasomnias occurring in NREM include bruxism (grinding teeth) and restless leg syndrome, in which the sleeper's legs jiggle (remember, there is no skeletal muscle paralysis in NREM). Parasomnias occurring in REM include REM sleep behavior disorder, in which muscle relaxation is incomplete, so that the person may act out some of the imagery in dreams (this is commonly observed in post-traumatic stress disorder). On the other hand, in recurrent isolated Sleep paralysis, the sleeper awakens with the muscle relaxation still in place, so that he cannot move (this dissipates relatively quickly). There is sleep apnea, pauses in breathing that can last for as little as a few seconds to several minutes, and which may occur as frequently as every two minutes or so. There's narcolepsy, in which people go from waking straight into REM sleep without passing through NREM -- often collapsing in the process, due to muscle relaxation.
|
Evans' studies of sleep suggestion suggest that information from the external environment can be processed during sleep and be retained into subsequent REM periods the same night and even on later nights. Evans' research was inspired, in part, by hypnosis, in which the subject acts on various suggestions administered by the hypnotist, and he wondered if something similar might occur during sleep. Evans understood clearly that, despite the origins of its name (hypnos is Greek for sleep), hypnosis wasn't anything like sleep, either physiologically or behaviorally. Still, an early study by Lawrence LeShan (1942), a clinical psychologist, had suggested (sorry) that subjects could respond to suggestions presented during sleep. LeShan worked with a group of boys at a summer camp who were prone to nail-biting, and he played them a phonograph record containing the suggestion that "My fingernails taste terribly bitter" (applying a bitter-tasting liquid to the fingernails was a common treatment for nail-biters at the time). After 300 repetitions per night over 54 nights (!), 40% of the subjects who received the suggestion stopped biting their nails, compared to none in a control group. Unfortunately, LeShan was not able make use of the EEG to confirm that his subjects actually slept during the presentations. T.X. Barber (1959), also a distinguished hypnosis researcher, found that subjects showed increased responsiveness to hypnotic suggestions during descending Stage 1 sleep.
Evans' general procedure was to recruit subjects for a standard sleep study, taking place over two nights, but making no mention of suggestions. After subjects fell asleep, he administered various hypnotic-like suggestions, typically over an intercom: A pilot study by Cobb et al. (1965)
seemed to show that subjects responded to the cues when they were
presented during REM sleep (i.e., about 20% of the time), but not
when the same cues were presented in the morning, after
awakening. 20% may not seem high, but it's a lot higher
than zero, which is what we'd expect to happen.
A pilot study by Cobb et al. (1965)
seemed to show that subjects responded to the cues when they were
presented during REM sleep (i.e., about 20% of the time), but not
when the same cues were presented in the morning, after
awakening. 20% may not seem high, but it's a lot higher
than zero, which is what we'd expect to happen.
 A more thorough replication
and extension of the Cobb study by Evans et
al. (1969, 1970) appeared to confirm this
result, with about a 20% response rate.
Evans also noted a fairly long latency between
cue and response, as if the subjects had to
mobilize themselves to respond to the
cue. The response rate on the delayed
tests, 23%, was about the same as on the
immediate tests. However, the latency of
response was longer on the delayed tests (c.
35 seconds) than it was on the immediate test
(c. 19 seconds).
A more thorough replication
and extension of the Cobb study by Evans et
al. (1969, 1970) appeared to confirm this
result, with about a 20% response rate.
Evans also noted a fairly long latency between
cue and response, as if the subjects had to
mobilize themselves to respond to the
cue. The response rate on the delayed
tests, 23%, was about the same as on the
immediate tests. However, the latency of
response was longer on the delayed tests (c.
35 seconds) than it was on the immediate test
(c. 19 seconds).
Upon awakening the next morning, the subjects displayed little memory for events of the previous night. On direct interview, 10 of the 19 subjects reported some awareness of the experimenter's presence. While they could occasionally recall parts of the suggestions or cues, they typically did not recall responding. When given a word-association test designed to elicit the suggested responses to the cues in the waking state, subjects gave the appropriate verbal response (e.g., itch-nose), but the suggested behavioral response was elicited in only 6% of the cases.
Moreover, some subjects continued to respond to the cues the next night, and even when tested 5 months later -- but not while awake. This suggested that response to sleep suggestions was in some sense state-dependent -- that is, it seemed the cues had to be administered in the same state as the suggestion, in order to be effective.
 However, a carefully
controlled investigation by Perry et al.
(including Evans as a co-author), with blind
judges evaluating response to "dummy" as well
as "real" cues, casts doubt on these earlier
findings. What appeared to be specific
responses to suggestions administered during
REM sleep were probably artifacts of
spontaneous activity during sleep, interpreted
by the non-blind experimenter as responses to
suggestions.
However, a carefully
controlled investigation by Perry et al.
(including Evans as a co-author), with blind
judges evaluating response to "dummy" as well
as "real" cues, casts doubt on these earlier
findings. What appeared to be specific
responses to suggestions administered during
REM sleep were probably artifacts of
spontaneous activity during sleep, interpreted
by the non-blind experimenter as responses to
suggestions.
If dreams are so poorly
remembered, and subjects are not responsive to
suggestions administered during sleep, what
are the possibilities for sleep learning?
Sleep learning is a
long-standing fantasy. In Aldous
Huxley's dystopian novel Brave New World
(1932), set in the 26th century, Bernard Marx,
a psychologist at the Central London Hatchery
and Conditioning Centre, employs
sleep-learning to indoctrinate new citizens of
the World State into the official ideology of
"Fordism", and acceptance of their assignment
within a hierarchical caste system. It's
a good plot device, but does that really work?
Perhaps the earliest study of sleep learning was conducted by Thorndike (1916), on military recruits learning Morse code (this was in the run-up to World War I). Records containing letter-Morse equivalents (e.g., A -- dit-dah; B -- dah-dit-dit-dit; C -- dah-dit-dah-dit) were played to the soldiers while they slept in their barracks. Thorndike reported that soldiers who heard the recordings while they slept learned code more quickly in their daytime classes; but, in a foreshadowing of things to come, he also reported that they were very fatigued the next day. Due to the circumstances of the experiment, Thorndike was not able to rule out the possibility that the soldiers in the experimental group had also engaged in surreptitious learning.
 A study by Fox and Robins
(1952), conducted in the context of the Cold
War (when the Soviet Union was also engaged in
sleep research), engaged students in the
learning of Chinese-English paired
associates. The pairs were presented to
the subjects during sleep, and then the
subjects were asked to learn the same pairs
the next morning. In a control
condition, the subjects were asked to learn incorrect
pairs. Compared to subjects who were
presented with music rather than paired
associates while they slept, subjects learned
the correct pairs more quickly, and incorrect
pairs more slowly, the next morning.
A study by Fox and Robins
(1952), conducted in the context of the Cold
War (when the Soviet Union was also engaged in
sleep research), engaged students in the
learning of Chinese-English paired
associates. The pairs were presented to
the subjects during sleep, and then the
subjects were asked to learn the same pairs
the next morning. In a control
condition, the subjects were asked to learn incorrect
pairs. Compared to subjects who were
presented with music rather than paired
associates while they slept, subjects learned
the correct pairs more quickly, and incorrect
pairs more slowly, the next morning.
 While these results were
promising, Simon & Emmons (1955, 1956)
criticized the studies of sleep learning then
available on the grounds that sleep was not
confirmed using EEG criteria (e.g.,
disappearance of alpha activity). In
formal experiments, they monitored EEG while
sleeping subjects were presented with new
information in question-and-answer format.
Subjects were instructed to report if they
heard anything during the night, and then
tested for recall and recognition of the
information in the morning. Reports of hearing
the material when presented at night, and
recalling and recognizing it in the morning,
all varied directly with the amount of alpha
activity in the EEG, with recognition falling
to chance levels in (alpha-free) Stage NREM.
While these results were
promising, Simon & Emmons (1955, 1956)
criticized the studies of sleep learning then
available on the grounds that sleep was not
confirmed using EEG criteria (e.g.,
disappearance of alpha activity). In
formal experiments, they monitored EEG while
sleeping subjects were presented with new
information in question-and-answer format.
Subjects were instructed to report if they
heard anything during the night, and then
tested for recall and recognition of the
information in the morning. Reports of hearing
the material when presented at night, and
recalling and recognizing it in the morning,
all varied directly with the amount of alpha
activity in the EEG, with recognition falling
to chance levels in (alpha-free) Stage NREM.
 They also found that
memory was best when alpha activity occurred
within 10 seconds of stimulus presentation.
They also found that
memory was best when alpha activity occurred
within 10 seconds of stimulus presentation.
These findings led Simon and Emmons (1955) to their now-famous conclusion:
Simon and Emmons' conclusion virtually killed research on sleep learning, except in the Soviet Union and Eastern Europe, where sleep was generally diagnosed by respiration and other signs of autonomic arousal rather than EEG criteria.
On the other hand, it is known from the encoding specificity principle that memory is best when the conditions of retrieval are congruent with the conditions of encoding. In line with observations of "state-dependent" or "context-dependent" memory induced by drug states, emotional states, and environmental settings, it is possible that dreams and sleep-learning episodes are encoded in secondary memory, but that access to them is state-dependent -- that is, we would remember dreams and other information very well if only retrieval were attempted in Stage REM! Unfortunately, state-dependency is not an easy hypothesis to test in the context of sleep, and the fate of Evans' studies of sleep suggestion suggest that neither our inability to remember many dreams nor the failure of sleep learning is due to state-dependent learning and memory.
Still, some kind of
context-dependency is suggested by studies by
Kenneth Paller, Delphine Oudiette, and
their colleagues (summarized in Scientific
American, 11/2018) on the role of sleep
in the consolidation of memory for material
learned prior to sleep (as in the Stickgold
and Walker studies). These investigators
had subjects learn to insert pictures (e.g.,
of a cat or a helicopter) into their correct
locations on a computer screen. During
learning, presentation of each picture was
accompanied by a related sound (like a "meow"
or the whirring of rotor blades). Then
the subjects were allowed to nap (a proper
nap, it turns out, enhances consolidation just
like a full-night's sleep does). But
while the subjects slept, the investigators
played the same sounds that had been presented
during the learning task (a procedure they
call Targeted Memory Reactivation, or TMR),
while the subjects were awake. Compared
to a control group that did not receive these
sounds, the experimental group showed better
post-sleep retention for the pictures'
locations. Later research indicated that
this improvement in memory consolidation was
correlated with the extent of spindle activity
during Stage 2. As with the studies of
Stickgold and Walker, the studies of Paller
and Oudiette indicate that a considerable
amount of "off-line" information processing
goes on during sleep, much of which promotes
memory consolidation.
One theory holds that sleep enhances memory consolidation by virtue of continued rehearsal -- as in the McNaughton rat study of place learning and the hippocampus. Playing the same sounds during sleep as had been presented during learning seems to promote this rehearsal, perhaps along the lines of the encoding-specificity principle (see the General Psychology lectures on "Memory"). At the very least, the effect indicates that sleepers can process information from the environment, while remaining asleep.
Returning to the problem of sleep-learning per se, some post-Simon and Emmons studies, employing standard EEG criteria for sleep, did find some evidence for sleep learning. However, in retrospect, these experiments employed testing procedures that could have drawn on implicit rather than explicit memory.
 Evans & Orchard (1969)
obtained some evidence for sleep learning in
alpha-free REM, with paired associates of the
form A IS FOR APPLE. Because the stimuli
capitalized on pre-existing letter-word
associations, the subjects cued-recall
performance could have been enhanced by
repetition or semantic priming.
Evans & Orchard (1969)
obtained some evidence for sleep learning in
alpha-free REM, with paired associates of the
form A IS FOR APPLE. Because the stimuli
capitalized on pre-existing letter-word
associations, the subjects cued-recall
performance could have been enhanced by
repetition or semantic priming.
 Similarly, Cooper and
Hoscovec (1972) obtained some evidence for
sleep learning of Russian-English paired
associates in alpha-free REM (however,
learning was much more efficient when the
items were presented during waking).
Again, paired-associate learning can always be
influenced by priming effects between the cue
and the target.
Similarly, Cooper and
Hoscovec (1972) obtained some evidence for
sleep learning of Russian-English paired
associates in alpha-free REM (however,
learning was much more efficient when the
items were presented during waking).
Again, paired-associate learning can always be
influenced by priming effects between the cue
and the target.
Like many of the successful studies from the Soviet Bloc, then, both the Evans & Orchard study and the Cooper & Hoscovec study employed procedures that, in retrospect, could have capitalized on implicit rather than explicit memory, raising the question of whether it might be possible to demonstrate sleep learning with tests of implicit memory.
 Wood et al. (1992)
performed just such a study, presenting two
types of paired associates (homophones +
disambiguating context, e.g., WAR-PEACE;
category + instance, e.g., METAL-GOLD) during
REM and NREM.
Wood et al. (1992)
performed just such a study, presenting two
types of paired associates (homophones +
disambiguating context, e.g., WAR-PEACE;
category + instance, e.g., METAL-GOLD) during
REM and NREM.
Each of the subjects spent three nights in the laboratory. Night 1 was an adaptation night (sleep is typically disrupted the first night that subjects spend in the laboratory). The other two nights were study nights. On each night, after falling asleep, the subjects were presented with paired associates via earphones -- on one night, homophone pairs; on the other, category pairs, counterbalanced across nights. For most of the subjects, the stimuli were presented during Stage REM; for some subjects, they were presented during Stage 2. Each list was presented for four or five times per night. Presentation of the stimulus lists was discontinued if the subject showed alpha activity in the EEG, or any other signs of arousal, such as body movements or SREMs. After each list presentation, the subjects were allowed to sleep for two minutes, and then awakened. A control group heard the lists while lying awake in a darkened room.
 On
On  tests involving homophone
spelling and category generation, sleeping
subjects showed no evidence of priming; by
contrast, priming effects were substantial in
waking control subjects. For the present, then
it appears that sleep learning does not occur,
whether measured by explicit or by implicit
memory.
tests involving homophone
spelling and category generation, sleeping
subjects showed no evidence of priming; by
contrast, priming effects were substantial in
waking control subjects. For the present, then
it appears that sleep learning does not occur,
whether measured by explicit or by implicit
memory.
Amazingly, the Wood et al. study remains the only published study to date that assessed sleep learning with tests of both explicit and implicit memory. But it seems definitive, why should we repeat it? Because it actually leaves a couple of interesting questions unanswered.
First, Wood's criteria for sleep, while rigorously responsive to the Simon-Emmons complaint, may have been too strict. Perhaps sleep learning would have occurred under more naturalistic conditions, in which there are transient arousals during the night. The subjects might still have displayed no learning on tests of explicit memory, but they might have showed some implicit memory. Something similar has been observed in studies of implicit memory for anesthesia. Explicit memory is abolished at BIS levels of 60 or less, while implicit memory is abolished at BIS levels below 20. But between those two values, anesthesia causes a profound impairment in explicit memory, while sparing implicit memory.
Speaking of anesthesia brings us to the second question. If general anesthesia spares implicit memory, why doesn't sleep? By any standard, the brain is more active during sleep, especially REM sleep, than it is in adequate anesthesia. Recall, however, that Wood et al. employed tests of semantic priming, while the successful experiments on anesthesia employed repetition priming. The sleeper may not have the cognitive resources available to perform the sorts of meaning analyses required to support semantic priming, but may well have had enough resources to perform the sorts of perceptual analyses required to support repetition priming.
So, we'd really like to see the Wood et al. experiment done again, with measures of repetition as well as semantic priming. If that experiment failed to yield positive results, we'd really be certain that, as Simon and Emmons put it, "Sleep learning is possible, to the extent that the subject stays awake".
One of the greatest puzzles of sleep concerns memory for dreams. We probably have, on average, at least 4-5 dreams every night, but we typically remember, at best, only one of these. The contrast between the large number of dreams that occur in the night, and the small number of dreams remembered in the morning, raises the question of what accounts for the remembering and forgetting of dreams.
Which takes us to perhaps the most puzzling thing about sleep: dreaming. It has been said of dreaming that "Every time we experience REM sleep, we literally go mad. By definition, psychosis is condition characterized by hallucinations and delusions. Dreaming, some sleep scientists say, is a psychotic state -- we fully believe that we see what is not there, and we accept that time, location, and people themselves can morph and disappear without warning. From ancient Greeks to Sigmund Freud to back-alley fortune-tellers, dreams have always been a source of enchantment and mystery...."Want to Fall Asleep? Read this Story" by Michael Finkel (National Geographic, 08/2018, p. 72).
This page last modified 11/02/2020.






