For background, see also the
General Psychology lectures on
"The Biological Basis of Mind and Behavior" |
 Mind and behavior can be
analyzed at any of three broad levels:
Mind and behavior can be
analyzed at any of three broad levels:
Traditionally, social psychology has served to link psychology with the other social sciences, such as sociology, anthropology, economics, and political science. And biological psychology has served to link psychology with the other biological sciences. With the advent of brain-imaging technologies such as PET and fMRI, psychology is now in a position to ask questions about how mental processes are implemented in the brain. This project is most advanced within cognitive psychology, with a plethora of studies of the neural bases of perception, memory, and the like. But social psychology, and especially social cognition, has been part of this scene as well. Hence the emergence of social neuroscience, also known as social-cognitive neuroscience, and, most recently, cultural neuroscience (Kitayama & Uskul, 2011).
The task of psychology is to understand the nature of human mental life, including the biological bases of mental life.
From a functionalist perspective,
psychologists try to understand the role that mental life plays
in adaptive behavior -- and, more broadly, the relation of mind
to other things (in James' phrase), including:
 In
the 19th century, William James (1842-1910) defined psychology
as the science of mental life, attempting to understand the
cognitive, emotional, and motivational processes that underlie
human experience, thought, and action. But even in the
earliest days of scientific psychology, there was a lively
debate about how to go about studying the mind.
Accordingly, there arose the first of several "schools" of
psychology, structuralism and functionalism.
In
the 19th century, William James (1842-1910) defined psychology
as the science of mental life, attempting to understand the
cognitive, emotional, and motivational processes that underlie
human experience, thought, and action. But even in the
earliest days of scientific psychology, there was a lively
debate about how to go about studying the mind.
Accordingly, there arose the first of several "schools" of
psychology, structuralism and functionalism.
One point of view, known as
structuralism tried to study mind in the abstract. Wundt
and others in this school argued that complex mental states were
assembled from elementary sensations, images, and
feelings. Structuralists used a technique known as
introspection (literally, "looking into" one's own mind) to
determine, for example, the elements of the experience of a
particular color or taste. The major proponents of
structuralism were
Another point of view, known as
functionalism, was skeptical of the structuralist claim that we
can understand mind in the abstract. Based on Charles
Darwin's (1809-1882) theory of evolution, which argued
that biological forms are adapted to their use, the
functionalists focused instead on what the mind does,
and how it works. While the structuralists emphasized the
analysis of complex mental contents into their constituent
elements, the functionalists were more interested in mental
operations and their behavioral consequences. Prominent
functionalists were:
The Two FunctionalismsPsychological functionalism is often called "Chicago functionalism", because its intellectual base was at the University of Chicago, where both Dewey and Angell were on the faculty (functionalism also prevailed at Columbia University). It is to be distinguished from the functionalist theories of mind associated with some modern approaches to artificial intelligence (e.g., the work of Daniel Dennett, a philosopher at Tufts University), which describe mental processes in terms of the logical and computational functions that relate sensory inputs to behavioral outputs. This philosophical functionalism, closely related to Skinnerian functional behaviorism, is quite different from the psychological "Chicago functionalism" of James, Dewey, and Angell. |
Social psychology, and in
particular social cognition, serves some of these purposes
well.
Until recently, however, social cognition, and social psychology more broadly, has pretty much ignored mind in body, the third leg in the functionalist triad. Most social-psychological experiments and theories stick to the psychological level of analysis, and make little or no reference to underlying biological processes. In this way, as in so many other ways, social cognition has followed the lead of cognitive psychology in general. Social cognition has been no exception to this rule.
But in the last few decades, and
especially since the 1990s, cognitive psychology (and, of
course, cognitive neuroscience) has made great advances in
understanding the biological basis of cognition and
behavior. Similarly, social psychology has become
increasingly interested in biology, as indicated by several
trends:
Contemporary social neuroscience has its origins in physiological psychology, a term coined by Wundt in the 1870s to refer to experimental (as opposed to speculative) psychology in all its forms. Later, the term physiological psychology came to cover expressly physiological studies of various aspects of sensory function and behavior -- especially animal studies.
The term neuropsychology emerged in the 1950s to cover experimental studies of mind in behavior in human neurological patients suffering from various forms of brain insult, injury, or disease. It appears that Karl Lashley was the first psychologist to refer to himself as a neuropsychologist (1955), and the journal Neuropsychologia was founded in 1963 for the express purpose of publishing human neuropsychological research.
 The term neuroscience was introduced
in 1963, referring to an expressly interdisciplinary effort to
study the nervous system in all its aspects, from the micro to
the macro levels of analysis.
The term neuroscience was introduced
in 1963, referring to an expressly interdisciplinary effort to
study the nervous system in all its aspects, from the micro to
the macro levels of analysis.
Initially, neuroscience consisted
of three subfields:
Link to a paper which traces the evolution of social neuroscience and related fields.
As a matter of personal preference, I prefer neuropsychology to neuroscience, because the former term places emphasis on the neural basis of mind and behavior, while the latter term places emphasis on -- well, neurons. Neuropsychology is a field of psychology, a discipline concerned with mind and behavior. Neuroscience is a field of biology, especially concerned with the anatomy and physiology of the nervous system. If you're interested in synapses and neurotransmitters, it makes sense to call yourself a neuroscientist. But if you're interested in the mind and behavior of the individual person, it makes more sense to identify yourself as a psychologist. But the term neuroscience is preferred by the majority of researchers in the field, incorporating neuropsychology as a particular subfield (focusing on behavioral studies of brain damage), and I'll bow to the inevitable.
 Whatever
Whatever  you call it,
neuropsychology or neuroscience, the field has its roots
in physiological psychology. In the 19th
century, physiological psychology worked in parallel with
psychophysics to put the study of mental life on
an objective basis -- physiological
psychology, by tying subjective mental states (like
sensations and percepts) to objectively observable nervous
system structures and processes; psychophysics by tying
those same subjective mental states to
objectively observable properties of physical
stimuli. But in the 20th century, physiological
psychology took on the task of identifying the relations between
behavior and physiological processes.
you call it,
neuropsychology or neuroscience, the field has its roots
in physiological psychology. In the 19th
century, physiological psychology worked in parallel with
psychophysics to put the study of mental life on
an objective basis -- physiological
psychology, by tying subjective mental states (like
sensations and percepts) to objectively observable nervous
system structures and processes; psychophysics by tying
those same subjective mental states to
objectively observable properties of physical
stimuli. But in the 20th century, physiological
psychology took on the task of identifying the relations between
behavior and physiological processes.
 This
is, essentially, the
same task that behavioral neuroscience (including
cognitive, social, affective, and conative
neuroscience) undertakes
today. Earlier, David Marr had proposed that cognition could be
analyzed at three quite different levels
(things always seem to go in threes!):
This
is, essentially, the
same task that behavioral neuroscience (including
cognitive, social, affective, and conative
neuroscience) undertakes
today. Earlier, David Marr had proposed that cognition could be
analyzed at three quite different levels
(things always seem to go in threes!):
So, to take an example from Palmer (1999), consider a thermostat:
Now,to take an example closer to social cognition, consider Anderson's "cognitive algebra" approach to impression-formation, as described in the lectures on Social Perception.
 Gazzaniga,
Gazzaniga,  in the very first statement of
the agenda for cognitive neuroscience, proposed to directly connect the algorithmic level to its
implementation in brain
tissue. In other words, to relate the mind's cognitive
functions to brain activity.
in the very first statement of
the agenda for cognitive neuroscience, proposed to directly connect the algorithmic level to its
implementation in brain
tissue. In other words, to relate the mind's cognitive
functions to brain activity.
Perhaps the simplest view of mind-brain relations is to think of the brain as a general-purpose information-processor, much like the central-processing unit of a high-speed computer. This is the view of the brain implicit in traditional learning theories, which assume that learning is a matter of forming associations between stimuli and responses. It is also the view implicit in traditional "empiricist" theories of the mind, which assume that knowledge and thought proceed by associations as well. Bits of knowledge are represented by clusters of neurons, and the associations between them are represented by synaptic connections -- or something like that.
 This was the view of
the brain which prevailed well into the 1960s, which
acknowledged relatively small areas of the brain devoted to
motor, somatosensory, visual, and auditory processing, but
construed the vast bulk of the brain as association cortex
-- devoted, as its name implies, to forming associations between
stimuli and responses.
This was the view of
the brain which prevailed well into the 1960s, which
acknowledged relatively small areas of the brain devoted to
motor, somatosensory, visual, and auditory processing, but
construed the vast bulk of the brain as association cortex
-- devoted, as its name implies, to forming associations between
stimuli and responses.
But if the brain were simply a general-purpose information-processing machine, there would be no reason to be talking about a distinctly cognitive or social neuropsychology or neuroscience, because there would be no reason to think that social cognition and behavior is performed any differently by the brain than is any other aspect of cognition and behavior.
A distinctly cognitive or social neuroscience is justified only by an alternative view, variously known as functional specialization, or localization of function, which holds that different psychological functions are performed by different parts of the brain. Thus, the brain is not a single, massive, undifferentiated organ, whose job it is to "think", but a collection of systems, each of which is dedicated to a particular cognitive, emotional, or motivational function. This viewpoint prevails in contemporary neuropsychology and neuroscience in general, as well as social neuropsychology and neuroscience in particular. Cognitive psychologists started taking the brain seriously in the late 1960s and early 1970s, but the whole idea of looking at the brain occurred to social psychologists only very recently (Klein & Kihlstrom, 1986).
 It is just this view which
underlies the Doctrine of Modularity, articulated by the
philosopher Jerry Fodor in 1983. According to Fodor, the
various input and output systems of the mind are structured as mental
modules. According to Fodor, these modules share a
number of properties in common, of which three are most
important for our purposes.
It is just this view which
underlies the Doctrine of Modularity, articulated by the
philosopher Jerry Fodor in 1983. According to Fodor, the
various input and output systems of the mind are structured as mental
modules. According to Fodor, these modules share a
number of properties in common, of which three are most
important for our purposes.
For a charming introduction to the application of the Doctrine of Modularity to neuroscience research, see "The Neuroanatomy Lesson (Director's Cut)" by Prof. Nancy Kanwisher at MIT.
 The
The  neuroscientific doctrine of
modularity traces its origins to Franz Joseph Gall (180-1919),
Spurzheim, and others who promoted a pseudoscience known as phrenology.
Phrenology was popularized in America by Nelson Sizer and O.S.
Fowler, and you can sometimes purchase a ceramic copy of
Fowler's "phrenological head" in novelty stores. Well into
the 19th century, the brain was thought to be a single, massive,
undifferentiated organ. But according to Gall and his
followers, the brain is composed of discrete organs, each
associated with some psychological faculty, such as morality or
love. In classic phrenological doctrine, there were about
35-40 such faculties, and the entire set of faculties comprises
a common-sense description of human abilities and
traits.
neuroscientific doctrine of
modularity traces its origins to Franz Joseph Gall (180-1919),
Spurzheim, and others who promoted a pseudoscience known as phrenology.
Phrenology was popularized in America by Nelson Sizer and O.S.
Fowler, and you can sometimes purchase a ceramic copy of
Fowler's "phrenological head" in novelty stores. Well into
the 19th century, the brain was thought to be a single, massive,
undifferentiated organ. But according to Gall and his
followers, the brain is composed of discrete organs, each
associated with some psychological faculty, such as morality or
love. In classic phrenological doctrine, there were about
35-40 such faculties, and the entire set of faculties comprises
a common-sense description of human abilities and
traits.
The Phrenological Faculties
From: "Phrenology,
the History of Brain Localization" |
 Gall and other
phrenologists inferred the associations between these abilities
and traits and brain locations by looking at exemplary cases:
if, for example, an obese person had a bulge in some part of his
skull, or a skinny person had a depression in the same area,
they reasoned that the area of the brain underneath must control
eating. In this way, the phrenologists inferred the
localization of each of the mental faculties in their system.
Gall and other
phrenologists inferred the associations between these abilities
and traits and brain locations by looking at exemplary cases:
if, for example, an obese person had a bulge in some part of his
skull, or a skinny person had a depression in the same area,
they reasoned that the area of the brain underneath must control
eating. In this way, the phrenologists inferred the
localization of each of the mental faculties in their system.
Gall's phrenological doctrines were embraced by August Comte, the 19th-century French philosopher who is generally regarded as the father of sociology. In his Positive Philosophy (Comte was also the father of positivism), he expressed general agreement with Gall's system, differing only in the number of faculties (which Comte called the "internal functions of the brain", or instincts. Comte accepted only 18 such functions, which he divided into three categories of Affective functions (located in the back or lower part of the brain), Intellectual functions (located in the front part of the brain), and Practical functions (located in the center of the brain). The following list is taken from an article by James Webb, reviewing the correspondence of Comte with John Stuart Mill (Phrenological Journal and Science, May 1900).
Comte was quite clear that each of these internal functions was mediated by a separate part of the brain. In fact, Comte founded a Church of Humanity as an enlightened, secular alternative to Christianity, with himself as the inaugural Grand Pontiff (he must have been quite ambitious, if not downright dotty), one of whose rituals was that "disciples should tap themselves each day three times on the back of the head, where the impulses of "Good Will", "Order", and "Progress" were stored" (Thomas Meaney, "The Religion of Science and Its High Priest" [review of August Comte: An Intellectual Biography by Mary Pickering], New York Review of Books (10/25/2012). According to Meaney, the last operational Temple of Humanity is located in Paris, at 5 Rue Payenne.
 Gall's basic idea
was pretty good,
Gall's basic idea
was pretty good,  but his science
was very bad (which is why we call phrenology a
pseudoscience). Nobody takes phrenology seriously these
days. However, evidence from neuropsychology
and neuroscience favors Gall's idea of functional
specialization, though not in Gall's extreme form. The
modern, scientifically based list of mental "faculties", and
corresponding brain regions, is much different from his.
but his science
was very bad (which is why we call phrenology a
pseudoscience). Nobody takes phrenology seriously these
days. However, evidence from neuropsychology
and neuroscience favors Gall's idea of functional
specialization, though not in Gall's extreme form. The
modern, scientifically based list of mental "faculties", and
corresponding brain regions, is much different from his.
 Comparing the
classical and modern phrenological heads, one thing stands out:
while modern neuropsychology and neuroscience have focused on
the brain structures and systems associated with nonsocial
cognition, such as color vision or the perception of motion, the
classical phrenologists were much more concerned with
personality and social functioning. Indeed, roughly half
of the classic phrenological faculties referred to personality
traits or social behaviors, as opposed to nonsocial aspects of
sensation, perception, memory, language, and reasoning.
Although social cognition and behavior was slighted in the early
years of neuropsychology and neuroscience, personality and
social psychology have begun to catch up.
Comparing the
classical and modern phrenological heads, one thing stands out:
while modern neuropsychology and neuroscience have focused on
the brain structures and systems associated with nonsocial
cognition, such as color vision or the perception of motion, the
classical phrenologists were much more concerned with
personality and social functioning. Indeed, roughly half
of the classic phrenological faculties referred to personality
traits or social behaviors, as opposed to nonsocial aspects of
sensation, perception, memory, language, and reasoning.
Although social cognition and behavior was slighted in the early
years of neuropsychology and neuroscience, personality and
social psychology have begun to catch up.
Phrenology was popular in the 19th century, but it was also very controversial. However, the doctrine of functional specialization gathered support from a number of case studies that appeared in the 19th century medical literature, which seemed to indicate that particular mental functions were performed by particular brain structures.
 Historically, perhaps the most
important of these studies concerned various forms of aphasia
-- a neurological syndrome involving the loss or impairment of
various speech and language functions. In neither case is
there any damage to the corresponding skeletal musculature: the
source of the psychological deficit resides in specific lesions
in the central nervous system.
Historically, perhaps the most
important of these studies concerned various forms of aphasia
-- a neurological syndrome involving the loss or impairment of
various speech and language functions. In neither case is
there any damage to the corresponding skeletal musculature: the
source of the psychological deficit resides in specific lesions
in the central nervous system.
 But
But  before Broca, and Wernicke, a similar point was made, or at
least debated, in the case of Phineas Gage reported by
John Martyn Harlow (1848, 1850, 1868), a physician who practiced
in rural Vermont in the mid-19th century. Gage was a
railway-construction foreman for the Rutland and Burlington
Railroad whose job it was to tamp blasting powder into
rock. At 4:30 PM on September 13, 1848 (a Wednesday), near
Duttonville (now Cavendish), Vermont, Gage accidentally set off
an explosion which drove his tamping iron through his left eye
socket and out the top of his head. Gage survived the
accident, was treated by Harlow, and after 12 weeks of
recuperation returned to his home in Lebanon, New
Hampshire.
before Broca, and Wernicke, a similar point was made, or at
least debated, in the case of Phineas Gage reported by
John Martyn Harlow (1848, 1850, 1868), a physician who practiced
in rural Vermont in the mid-19th century. Gage was a
railway-construction foreman for the Rutland and Burlington
Railroad whose job it was to tamp blasting powder into
rock. At 4:30 PM on September 13, 1848 (a Wednesday), near
Duttonville (now Cavendish), Vermont, Gage accidentally set off
an explosion which drove his tamping iron through his left eye
socket and out the top of his head. Gage survived the
accident, was treated by Harlow, and after 12 weeks of
recuperation returned to his home in Lebanon, New
Hampshire.
 Certainly
Certainly
 the most remarkable fact about the accident is that
Gage lived to tell about it. However, Harlow (1968) also
reported that Gage underwent a marked personality change at the
time of his accident.
the most remarkable fact about the accident is that
Gage lived to tell about it. However, Harlow (1968) also
reported that Gage underwent a marked personality change at the
time of his accident.
The Definitive Skinny on Gage
The mythology is
epitomized by an article by Hannah Damasio et al. (Science, 1994), which revived
interest in the Gage case in the
context of Anthony Damasio's theories of the role
of prefrontal cortex in rationality and emotion --
to which Gage's allegedly "profound personality
changes" (p. 1102), in which "Gage was no longer
Gage" (Harlow, 1868, p. 327) are relevant. .
Damasio's
paper made an important contribution to our
understanding of Gage by using skull measurements
(Gage's skull, though not his brain, is preserved
at Harvard Medical School) and modern
brain-imaging technology to reconstruct his
injury. Harlow, based on his physical
examination, had concluded that Gage's right
hemisphere was undamaged. Damasio et al. conclude that the
damage included the prfrontal cortex of both
hemispheres. But at the same time, they built
their account of Gage's behavior largely on
secondary sources,
which themselves are -- as
Macmillan has shown -- grossly exaggerate the
extent to which Gage had "taken leave of his
sense of responsibility" and no longer could
be "trusted to honor
his commitments" (p. 1102). All we really know about Gage from
first-hand medical examination comes from five
reports: three from Harlow (two of them written
close to the time of the
incident, the third written 20 years later), and two from Bigelow, both written years after
Gage's death. There is no question that Gage
was a responsible individual
before his accident.
He was a foreman, after all, and
railroads don't put high explosives
in the hands of irresponsible,
untrustworthy individuals. While it is
true that he lost his
position as foreman, he did attempt to return to work in
1849, but eventually began to suffer from epileptic
seizures presumably brought on by his brain damage.
It should be noted that most of the contemporary accounts of Gage's personality change, including Harlow's claim that "Gage was no longer Gage", were published years after his death. And, frankly, we don't know very much about Gage's pre-accident personality: he was just another man working on the railroadThe very first accounts, from Harlow (1948, 1849) and Bigelow (1950), focused mostly on the fact that Gage survived the accident -- although Harlow (1848) did refer to difficulties in controlling Gage. Later, his case was presented as evidence that specific brain damage did not cause specific loss of function -- evidence supporting attacks on phrenology and other claims about localization of function (Ferrier, 1876). Macmillan has uncovered evidence that Harlow himself was interested in phrenology, and it may well be that his reports of Gage's personality change were colored by his commitment to phrenological doctrine. It wasn't until 1878 that Ferrier himself, in his Gulstonian Lectures to the Royal College of Physicians, offered Gage as evidence favoring localization -- based on evidence from Harlow's later, posthumous, reports. But even in Harlow's 1868 report, he noted that Gage was fond of pets, children, horses, and dogs. Gage's full story is told in Macmillan's book, An Odd Kind of Fame: Stories of Phineas Gage (MIT Press, 2000). Those who don't have time for the whole book will find a useful precis in an earlier paper: "A Wonderful Journey Through Skulls and Brains: The Travels of Mr. Gage's Tamping Iron" (Brain & Cognition, 5, 67-107, 1986).
See also:
Macmillan also maintains an excellent webpage devoted to Gage, which contains much interesting material not in the book: http://www.deakin.edu.au/hbs/GAGEPAGE/. |
 According
According
 to some interpretations, Gage's
case was consistent with phrenology, because his damage seemed
to occur in the area of the frontal lobes associated with the
faculties of Veneration and Benevolence. Others disputed
the exact site of the damage, and others the magnitude of the
personality change. Subsequently, Gage became a linchpin
of arguments about phrenology in particular and functional
specialization in general. There is no question that Gage was injured, and that he subsequently
underwent a personality change. But Macmillan shows that
most popular and textbook accounts of his personality change
are not corroborated by the
actual evidence reported by Harlow and
others at the time. And, Macmillan discovered,
Harlow himself was a kind of closeted phrenologist, so
his representations and interpretations of Gage's personality changes may well have been
biased and colored by his theoretical commitments.
to some interpretations, Gage's
case was consistent with phrenology, because his damage seemed
to occur in the area of the frontal lobes associated with the
faculties of Veneration and Benevolence. Others disputed
the exact site of the damage, and others the magnitude of the
personality change. Subsequently, Gage became a linchpin
of arguments about phrenology in particular and functional
specialization in general. There is no question that Gage was injured, and that he subsequently
underwent a personality change. But Macmillan shows that
most popular and textbook accounts of his personality change
are not corroborated by the
actual evidence reported by Harlow and
others at the time. And, Macmillan discovered,
Harlow himself was a kind of closeted phrenologist, so
his representations and interpretations of Gage's personality changes may well have been
biased and colored by his theoretical commitments.
Gage and Lobotomy A common
interpretation of Gage is that he suffered the
accidental equivalent of a prefrontal lobotomy, and
it is sometimes held that Gage inspired the
development of psychosurgery in the 1930s
and 1940s, which (in those days before psychotropic
drugs) often sought to make psychiatric patients
more controllable by destroying portions of their
frontal lobes.
Malcolm Macmillan's historiography of Gage (see below) shows pretty conclusively that this was not the case. In the first place, the "new" Gage's symptoms were precisely the opposite of what prefrontal lobotomy was supposed to accomplish. There are many film
dramatizations of prefrontal lobotomies, including:
For definitive
histories of lobotomy and other forms of
psychosurgery, see two books by Eliot Valenstein
(himself a distinguished neuroscientist):
|
 The fact is, we
simply know too little about Gage's personality, and his injury,
to draw any firm conclusions about personality, social behavior,
and the brain. In the final analysis, the Gage case is
only suggestive of what was to come. But just as Broca's
patient Tan and Scoville and Milner's patient H.M. became the
index cases for cognitive
neuroscience, Phineas Gage deserves a place as the index case
for what has become social neuroscience.
The fact is, we
simply know too little about Gage's personality, and his injury,
to draw any firm conclusions about personality, social behavior,
and the brain. In the final analysis, the Gage case is
only suggestive of what was to come. But just as Broca's
patient Tan and Scoville and Milner's patient H.M. became the
index cases for cognitive
neuroscience, Phineas Gage deserves a place as the index case
for what has become social neuroscience.
 Interestingly, and
perhaps somewhat prophetically, Gage is an ancestor of Fred
Gage, a Salk Institute neuroscientist who studies
neurogenesis. After Smithsonian
Magazine published a newly
discovered photograph of Gage (01/2010),
two of his other descendants
independently contributed yet another
image of him with his tamping-iron (published in the magazine in
03/2010). The original image was a
daguerreotype, or something like it, which is a mirror image of the real
object. Here, the image has been rotated
vertically, to correctly show that Gage's injury was to his left eye.
Interestingly, and
perhaps somewhat prophetically, Gage is an ancestor of Fred
Gage, a Salk Institute neuroscientist who studies
neurogenesis. After Smithsonian
Magazine published a newly
discovered photograph of Gage (01/2010),
two of his other descendants
independently contributed yet another
image of him with his tamping-iron (published in the magazine in
03/2010). The original image was a
daguerreotype, or something like it, which is a mirror image of the real
object. Here, the image has been rotated
vertically, to correctly show that Gage's injury was to his left eye.
 So given the hypotheses the
brain damage has consequences for social cognition, and that
there are brain systems that are specialized for
social-cognitive tasks, how would we know? Neuropsychology
offers a number of techniques for studying functional
specialization.
So given the hypotheses the
brain damage has consequences for social cognition, and that
there are brain systems that are specialized for
social-cognitive tasks, how would we know? Neuropsychology
offers a number of techniques for studying functional
specialization.
Historically, the most important
method for neuropsychology involves neurological cases of
patients with brain lesions, in which scientists observe
the mental and behavioral consequences of damage to some portion
of the brain.
Here are some combined MRI and PET images of the brain of a single subject engaged in different kinds of thinking, collected by Dr. Hanna Damasio of the University of Iowa College of Medicine, and published in the New York Times Magazine (May 7, 2000). Red areas indicate increased brain activation, as indicated by blood flow, while areas in purple indicate decreased brain activation.
- In the upper left corner, the subject is thinking pleasant thoughts;
- in the upper right corner, depressing thoughts.
- In the lower left corner, anxious thoughts;
- in the lower right, irritating thoughts.
Brain imaging techniques are
gaining in popularity, especially in human research, because
they allow us to study brain-behavior relations in human
subjects who have intact brains. But still, some of the
clearest evidence for mind-brain relations is based on the
lesion studies that comprise classic neuropsychological
research.

 The Case of H.M. As an illustration of
the role that neuropsychological evidence from brain-damaged
patients can play in understanding functional specialization,
consider the case of a patient, known to science by the
initials H.M., who suffered from a severe case of epilepsy
involving frequent, uncontrollable seizures that seemed to arise
from his temporal lobes. As an act of desperation, H.M.
agreed to surgical excision of the medial (interior) portions of
his temporal lobe -- an operation that also excised his hippocampus
and portions of the limbic system, including the
hippocampus. After surgery, he behaved normally with
respect to locomotion, emotional responses, and social
interaction -- except that he had no memory. H.M.
displayed a retrograde amnesia for events occurring
during the three years prior to the surgery. But more
important, he displayed an anterograde amnesia for all
new experiences. He could not remember current events and
he could not learn new facts. The retrograde amnesia
remitted somewhat after the surgery, but the anterograde amnesia
persists to this day. As of 2003, H.M. was still alive,
but he remembers nothing of what has happened to him since the
day of his surgery in 1953. He reads the same magazines,
and works on the same puzzles, day after day, with no
recognition that they are familiar. He meets new people,
but when he meets them again he does not recognize them as
familiar.
The Case of H.M. As an illustration of
the role that neuropsychological evidence from brain-damaged
patients can play in understanding functional specialization,
consider the case of a patient, known to science by the
initials H.M., who suffered from a severe case of epilepsy
involving frequent, uncontrollable seizures that seemed to arise
from his temporal lobes. As an act of desperation, H.M.
agreed to surgical excision of the medial (interior) portions of
his temporal lobe -- an operation that also excised his hippocampus
and portions of the limbic system, including the
hippocampus. After surgery, he behaved normally with
respect to locomotion, emotional responses, and social
interaction -- except that he had no memory. H.M.
displayed a retrograde amnesia for events occurring
during the three years prior to the surgery. But more
important, he displayed an anterograde amnesia for all
new experiences. He could not remember current events and
he could not learn new facts. The retrograde amnesia
remitted somewhat after the surgery, but the anterograde amnesia
persists to this day. As of 2003, H.M. was still alive,
but he remembers nothing of what has happened to him since the
day of his surgery in 1953. He reads the same magazines,
and works on the same puzzles, day after day, with no
recognition that they are familiar. He meets new people,
but when he meets them again he does not recognize them as
familiar.
 Work with H.M. and
similar patients reveals a brain system important for
memory. This circuit, known as the medial
temporal-lobe memory system, includes the hippocampus and
surrounding structures in the medial portion of the temporal
lobe. This circuit is not where new memories are
stored. But it is important for encoding new memories so
that they can be retrieved later. H.M. lacks this circuit,
and so he remembers nothing of his past since the surgery.
Work with H.M. and
similar patients reveals a brain system important for
memory. This circuit, known as the medial
temporal-lobe memory system, includes the hippocampus and
surrounding structures in the medial portion of the temporal
lobe. This circuit is not where new memories are
stored. But it is important for encoding new memories so
that they can be retrieved later. H.M. lacks this circuit,
and so he remembers nothing of his past since the surgery.
For an insightful portrayal of H.M.'s life, read Memory's Ghost by Philip J. Hilts.
 The
Case of S.M. Another example comes from the patient
S.M., who suffered damage to the amygdala, another
subcortical structure, but no damage to the hippocampus and
other structures associated with the medial temporal
lobes. S.M. suffers no memory deficit, nor any other
problems in intellectual functioning (Adolphs, Tranel, Damasio,
& Damasio, 1994). But he does display gross deficits
in emotional functioning: a general loss of emotional
responses to events; an inability to recognize facial
expressions of emotion; and an inability to produce appropriate
facial expressions himself. Research on S.M. and similar
patients suggests that the amygdala is part of another brain
circuit that is important for regulating our emotional life,
particularly fear (LeDoux).
The
Case of S.M. Another example comes from the patient
S.M., who suffered damage to the amygdala, another
subcortical structure, but no damage to the hippocampus and
other structures associated with the medial temporal
lobes. S.M. suffers no memory deficit, nor any other
problems in intellectual functioning (Adolphs, Tranel, Damasio,
& Damasio, 1994). But he does display gross deficits
in emotional functioning: a general loss of emotional
responses to events; an inability to recognize facial
expressions of emotion; and an inability to produce appropriate
facial expressions himself. Research on S.M. and similar
patients suggests that the amygdala is part of another brain
circuit that is important for regulating our emotional life,
particularly fear (LeDoux).
The contrast between patients H.M. and S.M. illustrate the logic of dissociation that is central to cognitive neuropsychology. By "dissociation" we simply mean that some form of brain damage (or, for that matter, any other independent variable) affects one aspect of task performance but not another. Thus, "dissociation" is analogous to the interaction term in the statistical analysis of variance. In the case of H.M., damage to the hippocampus impairs performance on memory but not emotional tasks; but in the case of S.M., damage to the amygdala impairs performance on emotional tasks but not memory. Therefore, we can conclude that the hippocampus is part of a brain system that is specialized for memory, while the amygdala is part of a separate (dissociable) brain system that is specialized for emotion. Of course, this assumes that the experimenter has eliminated potential experimental confounds.
The fact that the nervous system operates according to certain electrochemical principles has opened up a wide range of new techniques for examining the relationship between brain and mind. Some of these techniques are able to detect lesions in the brain, without need for exploratory surgery or autopsy. Others permit us to watch the actual activity of the brain while the subject performs some mental function.
CT (CAT) Scans. In x-ray computed tomography (otherwise known as CAT scan, or simply CT), x-rays are used to produce images of brain structures. This would seem to be an obvious application of x-ray technique, but there are some subtle problems: (1) radiation can damage brain tissue; (2) brain tissue is soft, and so x-rays pass right through it; and (3) x-rays produce two-dimensional images, and so it is hard to distinguish between overlapping structures (that is, you can see the edges of the structures, but you can't detect the boundary between them). The CT scan uses extremely low doses of x-rays, too weak to do any damage, or to pass through soft tissue. It also takes many two-dimensional images of the brain, each from a different angle. Then a computer program takes these hundreds of individual two-dimensional images and reconstructs a three-dimensional image (this requires a very fast, very powerful computer). CT scans allow us to determine which structures are damaged without doing surgery, or waiting for the patient to die so that an autopsy can be performed.
Magnetic Resonance Imaging (MRI). The technique of magnetic-resonance imaging (MRI) is based on the fact that some atoms, including hydrogen atoms, act like tiny magnets: when placed in a magnetic field, they will align themselves along lines of magnetic force. Bombarding these atoms with radio waves will set them spinning, inducing a magnetic field that can be detected by sensitive instruments. In a manner similar to CT, readings from these instruments can be used to reconstruct a three-dimensional image of the brain. However, this image has a much higher resolution than CT, and so can detect much smaller lesions.
MRI is such an important advance in medical technology that Nobel prizes have been awarded on several occasions for work relating to it. The 2003 Nobel Prize for Medicine or Physiology was awarded to Paul C. Lauterbur, a physical chemist at the University of Illinois, and Peter Mansfield, a physicist at the University of Nottingham, in England, for basic research that led to the development of the MRI. Lauterbur published a pioneering paper on 2-dimensional spatial imaging with nuclear magnetic resonance spectroscopy (when NMR was picked up by medicine, the word "nuclear" was dropped for public-relations reasons, so that patients would not think that the technique involved radiation -- which it doesn't). Mansfield later developed a technique for 3-dimensional scanning. Eager young scientists should note that Lauterbur's paper was originally rejected by Nature, although the journal eventually published a revision. And if that's not inspiration enough, Mansfield dropped out of the British school system at age 15, returning to college later; now he's been knighted!
Some controversy ensued because the prize committee chose not to honor the contributions of Raymond Damadian, a physician, inventor, and entrepreneur, who made the initial discovery that cancerous tissue and normal tissue give off different magnetic resonance signals. Damadian also proposed that it be used for scanning tissues inside the body, and made the first working model of a MR scanner (now on display in the Smithsonian National Museum of American History). Damadian subsequently took out expensive full-page advertisements in the New York Times and other publications to assert his priority. But even before Damadian, Vsevolod Kudravcev, an engineer working at the National Institutes of Health, produced a working MRI device in the late 1950s by connecting an NMR to a television set: his supervisor told him to get back to his assigned job, and nothing came of his work. (See "Prize Fight" by Richard Monastersky, Chronicle of Higher Education, 11/07/03).
The Nobel committee, following its tradition, has been silent on its reasons for excluding Damadian. Everyone seems to agree that Lauterbur and Mansfield's work was crucial to developing MRI as a clinically useful technique. Still, no one doubts the importance of Damadian's pioneering work, either, and the Nobel rules make room for up to three recipients of a prize. The decision may just reflect an admittedly unreliable historical judgment. On the other hand, science has its political elements, and there could have been other reasons for denying Damadian the prize. It is possible that Damadian was denied the prize because he is a practicing physician and business entrepreneur rather than an academic scientist. Perhaps because he is relentlessly self-promoting (as in his newspaper as, which are without precedent in Nobel history), and famously litigious (he won a settlement of $129 million from General Electric for patent infringement) and rubs people the wrong way. Perhaps the Nobel committee did not want to give an award for biology, and thus at least indirect legitimacy, to someone who rejects the theory of evolution, the fundamental doctrine of modern biology, believes in a literal reading of the Bible, and has lent his support to creationism.
Positron Emission Tomography (PET). CT and MRI are new ways of doing neuroanatomy: they help us to localize brain damage without resort to surgery or autopsy. And that's important. But we'd also like to be able to do neurophysiology in a new way: to watch the brain in action during some mental operation. A technique that permits this is positron-emission tomography (PET). This technique is based on the fact that brain activity metabolizes glucose, or blood sugar. A harmless radioactive isotope is injected into the bloodstream, which "labels" the glucose. This isotope is unstable, and releases subatomic particles called positrons; the positrons collide with other subatomic particles called electrons, emitting gamma rays that pass through the skull and, again, are detected by sensitive instruments. When a particular portion of the brain is active, it metabolizes glucose, and so that part of the brain emits more gamma rays than other parts. Fast, powerful computers keep track of the gamma-ray emissions, and paint a very pretty picture of the brain in action, with different colors reflecting different levels of activity.
Functional MRI (fMRI). This is a variant on MRI, but with a much shorter temporal resolution than standard MRI. Like PET, it can be used to record the activity of small regions of the brain over relatively small intervals of time. However, the temporal resolution of fMRI is smaller than that of PET, permitting investigators to observe the working brain over shorter time scales. Whereas most brain-imaging studies have to "borrow" time on machines intended for clinical use, UC Berkeley has a very powerful fMRI machine dedicated entirely to research purposes.
Event-Related Potentials (Evoked Potentials). As you can imagine, CT, MRI, and PET are all very expensive, and require lots of equipment (in the case of PET, for example, a small nuclear reactor to produce the radioactive isotope). A very promising technique that does not have these requirements is based on the electroencephalogram (EEG), and is known as event-related potentials (ERPs, sometimes known as evoked potentials). In ERP, as in conventional EEG, electrodes are placed on the scalp to record the electrical activity of the neural structures underneath. Then, a particular stimulus is presented to the subject, and the response in the EEG is recorded. If you present the stimulus just once, you don't see much: there are lots of neurons, and so there's lots of noise. But in ERP, the brain's response to the same (or very similar) stimulus is recorded over and over again: when all the responses are combined, a particular waveform appears that represents the brain's particular response to that particular kind of event. The ERP has several components. Those that lie in the first 10 milliseconds or so reflect the activity of the brainstem; those that lie in the next 90 milliseconds, up to 100 milliseconds after the stimulus, reflect the activity of sensory-perceptual mechanisms located in the primary sensory projection area corresponding to the modality of the stimulus (seeing, hearing, touch, smell, etc.); those that lie beyond 100 milliseconds reflect the activity of cortical association areas. Interestingly, the characteristics of these components varies with the nature of the subject's mental activity. For example, the N100 wave, a negative potential occurring about 100 milliseconds after the stimulus, increases if the stimulus was in the focus of the person's attention, and decreases if it were in the periphery. The N200 wave, another negative potential appearing 200 milliseconds after the stimulus, is elicited by events that violate the subject's expectations. The P300 wave, a positive potential about 300 milliseconds out, is increased by some unexpected, task-relevant event (such as a change in the category to which the stimulus belongs); it seems to reflect a sort of "updating" of the subject's mental model of the environment. And the N400 wave, a negative potential about 400 milliseconds after the stimulus, is increased by semantic incongruity: for example, when the subject hears a nonsensical sentence.
ERPs can be recorded from the scalp as a whole, or they can be collected individually at lots of separate sites. In the latter case, the availability of powerful, high-speed computers permits a kind of brain-imaging: we can see where the ERP changes are the largest (or the smallest); and we can see how the ERP changes move with time. In this way, we can see how the brain shifts from one area to another when processing a stimulus or performing a task.
TMS. CAT, PET, MRI, and RP are all non-invasive, passive, recording technologies: that is, they employ sensors on the outside of the skin to record activity that naturally occurs under the skin (inside the skull, to be exact) when subjects are engaged in various tasks. Transcranial magnetic stimulation (TMS) is different, because it creates the functional equivalent of a temporary, reversible lesion in a discrete portion of brain tissue without requiring surgery to open up the scalp and skull (other techniques for creating temporary, reversible lesions, such as hypothermia and spreading depression, require surgical access to brain tissue). In TMS, a magnetic coil is applied to the scalp, and a magnetic pulse is delivered. This pulse can approach the magnitude of 2 Tesla, about the strength of the MRIs used clinically, but not as strong as the pulses produced by the machines used for research at UC Berkeley. The rapidly changing magnetic field induces an electrical field on the surface of the cortex. This field in turn generates neural activity which is superimposed on, and interferes with, the ongoing electrical activity of nearby portions of the brain. This temporary disruption of cortical activity, then, interferes with the performance of tasks mediated by parts of the brain near the site of application. For example, TMS applied over a particular region of the occipital lobe interferes with visual imagery, supporting findings from other brain-imaging techniques that striate cortex is involved in visual imagery, as it is in visual perception. TMS is useful because it has better temporal resolution, and equal spatial resolution, than other available techniques, such as PET and fMRI. That is, it can record the activity of relatively small areas over very small units of time.
Whatever the method, brain-imaging data is analyzed by some variant of the method of subtraction. Brain images are generated while subjects perform a critical task, and then while performing a control task. The pattern of activation associated with the control task is subtracted from the pattern associated with the critical task, leaving as a residue the pattern of brain activation that is specifically associated with the critical task. Of course, the success of the method depends on the tightness of the experimental controls. To the extent that the comparison between critical and control tasks is confounded by uncontrolled variables, the pattern of activation associated with the critical task may well be artifactual.
 The application of the idea of
modularity to social cognition and behavior was anticipated in
the theory of multiple intelligences proposed by Howard
Gardner (1983), a cognitive psychologist at Harvard -- and
author of The Mind's New Science (Basic Books, 1985), an
excellent account of the cognitive revolution in psychology and
the founding of interdisciplinary cognitive science. The
theory is proposed in the context of the debate over the
structure of human intelligence. According to Gardner,
intelligence is not a single ability, such as the g (or
"general intelligence") favored by Spearman and Jensen, but
rather a collection of abilities, much like the primary
mental abilities of Thurstone or Guilford's structure
of intellect. The system of multiple intelligences
proposed by Gardner consists of seven separate abilities:
The application of the idea of
modularity to social cognition and behavior was anticipated in
the theory of multiple intelligences proposed by Howard
Gardner (1983), a cognitive psychologist at Harvard -- and
author of The Mind's New Science (Basic Books, 1985), an
excellent account of the cognitive revolution in psychology and
the founding of interdisciplinary cognitive science. The
theory is proposed in the context of the debate over the
structure of human intelligence. According to Gardner,
intelligence is not a single ability, such as the g (or
"general intelligence") favored by Spearman and Jensen, but
rather a collection of abilities, much like the primary
mental abilities of Thurstone or Guilford's structure
of intellect. The system of multiple intelligences
proposed by Gardner consists of seven separate abilities:
Gardner's theory is a psychological theory of intelligence, not a theory of the neural correlates of intelligence. However, because he employs neurological evidence to bolster his case for multiple, independent intelligences, it implies that certain aspects of personality and social interaction -- including, presumably, certain aspects of social cognition -- are performed by specialized brain systems.
Based mostly on animal research, Gray (1972, 1981, 1982, 1987) argued that there are two basic dimensions of personality, anxiety and impulsiveness, which have their biological bases in three separate and independent brain systems.
Gray's "biopsychological" system, in turn,
laid the foundation for Carver and Schier's
(1986, 1998, 2002) cybernetic control theory approach to self-regulation, which is
built on the interactions between BAS and
BIS (FFS having been left by the wayside) -- as well as many
other theories.
Carver and White (1994) developed
questionnares for the measurement of BAS and BIS activity, while
Jackson (2009) developed alternative scales for BIS and BAS, and
added scales for the three different aspects of FFFS.
In a discussion of the neural
substrates of social
intelligence, Taylor and Cadet (1989) proposed that there
are at least three different social brain subsystems:
 As part of an
influential theory of childhood autism, to be discussed in the
lectures on Social-Cognitive
Development, Baron-Cohen (Simon, that is, cousin of Sacha,
the comedian) has proposed in 1995 that there are four elements of
mindreading -- each organized as a mental module, and each
associated with a different neural substrate.
As part of an
influential theory of childhood autism, to be discussed in the
lectures on Social-Cognitive
Development, Baron-Cohen (Simon, that is, cousin of Sacha,
the comedian) has proposed in 1995 that there are four elements of
mindreading -- each organized as a mental module, and each
associated with a different neural substrate. And in a popular
treatment of the literature on social intelligence,
Daniel Goleman ((2006) proposed several functions of the social
brain, each presumably modular in nature:
And in a popular
treatment of the literature on social intelligence,
Daniel Goleman ((2006) proposed several functions of the social
brain, each presumably modular in nature: The most extensive proposal concerning social-cognitive modules has come from
Ray Jackendoff (1992, 1994), a linguist and cognitive
scientist at Brandeis University, who was directly influenced by
Fodor. While Fodor was
concerned only with mental modules for input and output
functions associated with vision and language, he left room for
central modules as well, and Jackendoff proposed a number of candidates:
The most extensive proposal concerning social-cognitive modules has come from
Ray Jackendoff (1992, 1994), a linguist and cognitive
scientist at Brandeis University, who was directly influenced by
Fodor. While Fodor was
concerned only with mental modules for input and output
functions associated with vision and language, he left room for
central modules as well, and Jackendoff proposed a number of candidates:
Jackendoff's arguments for a
mental faculty of social cognition are mostly
philosophical in nature:
Everybody's familiar with Alzheimer's disease (AD), a chronic
neurodegenerative disease form of dementia mostly (though not
exclusively) associated with aging, and primarily affecting the
temporal and parietal lobes of the brain. The core symptoms
of AD are "cognitive" in nature, especially affecting short-term
as well as long-term memory -- which is why AD is often diagnosed
with tests of memory, and AD patients are commonly cared for in
facilities for "memory care".
But there's another, less-well-known form of dementia, known as Pick's disease or fronto-temporal dementia, where the degeneration primarily (as its name implies) affects the frontal and temporal lobes. Whereas AD is commonly diagnosed in the elderly, FTD is the most common form of dementia affecting people youner than 60. And while the primary symptoms of AD are cognitive in nature, the primary symptoms of FTD are social and emotional (Levenson & Miller, 2007). Call it a "social" dementia. FTD was first described by Arnold Pick, a Czech neurologist, in 1892, but it went largely unnoticed as medicine, science, and health policy focused on the challenge of AD.
The characteristics symptoms of FTD are frequently mistaken for those of depression - -or even of "midlife crisis".:
 In terms
of neuropathology, FTD appears to be associated with loss of
large, spindle-shaped "von Economy neurons" which congregate in
the anterior cingulate cortex, frontal portion of the insula, the
frontal pole, orbitofrontal cortex, and temporal pole. At
the cellular level, FTD seems to involve a build-up of tau
proteins, but not the beta-amyloid that is characteristically
seen, along with tau, in AD.
In terms
of neuropathology, FTD appears to be associated with loss of
large, spindle-shaped "von Economy neurons" which congregate in
the anterior cingulate cortex, frontal portion of the insula, the
frontal pole, orbitofrontal cortex, and temporal pole. At
the cellular level, FTD seems to involve a build-up of tau
proteins, but not the beta-amyloid that is characteristically
seen, along with tau, in AD.
The suggestion is that these structures constitute a network that
forms the neural basis of personality and character.
Degeneration of the neurons in this network leads to a breakdown
in the individual's normal personality -- with the specific
aspects lost depending on the site of the most serious
damage. Of course, these same centers are involved in
various cognitive functions, such as judgment and decision-making
(JDM), raising questions about whether FTD is specifically related
to personality. Then again, from a cognitive point of view,
personality and JDM are closely related.
Many of the leading experts on FTD are associated with the UC, including Profs. Robert Levenson and Robert Knight at UC Berkeley, and Profs. Bruce Miller and William Seeley at UCSF.
All of these proposals are more or less abstract, in which their authors have suggested that certain social-cognitive functions are modular in nature, without taking the next step to indicate where the modules are. However, brain-imaging research has begun to identify brain modules, or systems, associated with particular social-cognitive functions.
 Early
in
the development of social neuroscience, for example, Nancy
Kanwisher and her colleagues claimed to have identified three
such modules:
Early
in
the development of social neuroscience, for example, Nancy
Kanwisher and her colleagues claimed to have identified three
such modules:
 By
2007, Lieberman had identified more than 20 discrete brain areas
that are activated when subjects perform various social-cognitive
tasks. Based on spatial proximity in the brain, he
classified these modules into one of four categories based on two
core processes.:
By
2007, Lieberman had identified more than 20 discrete brain areas
that are activated when subjects perform various social-cognitive
tasks. Based on spatial proximity in the brain, he
classified these modules into one of four categories based on two
core processes.:However, identification of brain areas associated with various social-cognitive functions is only as good as the experimental paradigms used to tap those functions. This is not a trivial matter, as illustrated by certain areas of research.
You will remember, from the lectures on The Self, that neuropsychological data, especially from amnesia patients, shed light on the self as a memory structure. As summarized by Klein & Kihlstrom (1998), for example, it appears that amnesic patients retain accurate knowledge of their personality characteristics (semantic self-knowledge), even though they do not remember events and experiences in which they displayed those characteristics (episodic self-knowledge).
More recently, a variety of investigators have used brain-imaging technologies so search for a module specifically dedicated to processing self-relevant information.
In a pioneering experiment, Craik et al. (1999) employed PET to explore the neural correlates of the self-reference effect (SRE) in memory.
The SRE was originally uncovered
by Rogers and his colleagues, in a variant on the levels of
processing (LOP) paradigm for the study of human memory
(Craik & Lockhart). In a typical LOP experiment,
subjects are asked to make various judgments concerning words:
Rogers et al. employed trait
adjectives as their stimulus materials. More important,
they added a fourth condition to the standard LOP paradigm:
 On
the memory test, Rogers et al. replicated the standard LOP
effect. But more important, the self-referent orienting task
produced an even greater increment in memory. Based
on the standard interpretation of the LOP, Rogers et al. concluded
that the self is a very rich, elaborate memory structure --
perhaps the richest, most elaborate memory structure of all.
On
the memory test, Rogers et al. replicated the standard LOP
effect. But more important, the self-referent orienting task
produced an even greater increment in memory. Based
on the standard interpretation of the LOP, Rogers et al. concluded
that the self is a very rich, elaborate memory structure --
perhaps the richest, most elaborate memory structure of all.
Inspired by the SRE, Craik et al.
(1999) presented their subjects, who were Canadian college
students, with trait adjectives and scanned their brains while
they made one of four decisions about each word:
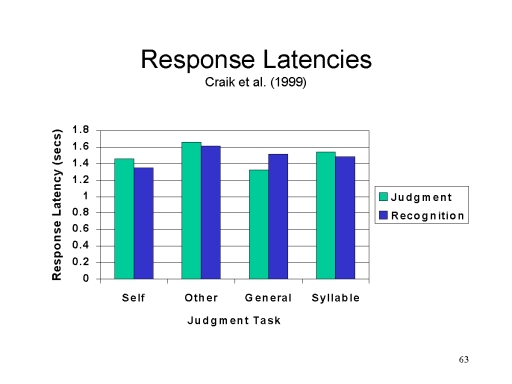 Because
brain activity varies with task difficulty, it was important to
show that the tasks were roughly equal in that respect, as
measured by response latencies.
Because
brain activity varies with task difficulty, it was important to
show that the tasks were roughly equal in that respect, as
measured by response latencies.
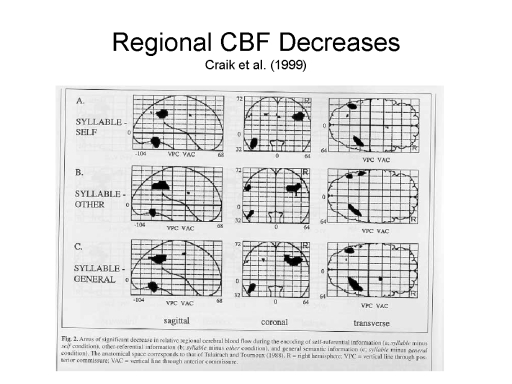 Using the subtraction
method, they then sought to
Using the subtraction
method, they then sought to  determine whether
there were any portions of the brain that were differentially
active in the self-referent condition, compared to the other
condition, as indicated by regional cerebral blood flow
(RCBF) measurements. In fact, there were some areas where
self-reference showed increases, and other areas where
self-reference showed decreases, compared to at least some
comparison conditions. But the most important
comparison was between the self-referent and other-referent
conditions.
determine whether
there were any portions of the brain that were differentially
active in the self-referent condition, compared to the other
condition, as indicated by regional cerebral blood flow
(RCBF) measurements. In fact, there were some areas where
self-reference showed increases, and other areas where
self-reference showed decreases, compared to at least some
comparison conditions. But the most important
comparison was between the self-referent and other-referent
conditions.
 The general area of
activation is indicated on the accompanying picture -- except that
this is a view of the left cerebral hemisphere, not the
right. Still you can get the idea of where the area is.
The general area of
activation is indicated on the accompanying picture -- except that
this is a view of the left cerebral hemisphere, not the
right. Still you can get the idea of where the area is.
 The Craik et al.
experiment was important as a pioneering effort to locate the
brain systems involved in self-referent processing, but it has
some problems. In particular, everything rests on the
comparison of self- and other-reference. In the Craik et
al. experiment, the target of the other-reference task was
probably not as familiar, or as well-liked, as the self.
In fact, as early as 1980 Keenan and Baillet had shown that the
SRE was matched by an other-reference task, provided
that the other person was well-known. Processing trait
adjectives with respect to a less-well-known other, such as
Jimmy Carter (for American subjects), produced no advantage
compared to standard semantic processing. The problem is
that the "other" in the Craik et al. experiment, Canadian Prime
Minister Brian Mulroney, was probably as unfamiliar to their
Canadian subjects as President Carter was to American
subjects. Put another way, Craik et al. might have gotten
quite different results if they had used a more appropriate
comparison condition -- one involving a highly familiar other
person.
The Craik et al.
experiment was important as a pioneering effort to locate the
brain systems involved in self-referent processing, but it has
some problems. In particular, everything rests on the
comparison of self- and other-reference. In the Craik et
al. experiment, the target of the other-reference task was
probably not as familiar, or as well-liked, as the self.
In fact, as early as 1980 Keenan and Baillet had shown that the
SRE was matched by an other-reference task, provided
that the other person was well-known. Processing trait
adjectives with respect to a less-well-known other, such as
Jimmy Carter (for American subjects), produced no advantage
compared to standard semantic processing. The problem is
that the "other" in the Craik et al. experiment, Canadian Prime
Minister Brian Mulroney, was probably as unfamiliar to their
Canadian subjects as President Carter was to American
subjects. Put another way, Craik et al. might have gotten
quite different results if they had used a more appropriate
comparison condition -- one involving a highly familiar other
person.
 Ochsner, Beer, and their colleagues (full
disclosure: I was one of them!) reported just such a study in
2005, in which self-referent judgments were compared with
judgments involving the subjects' best friends (there
were also two control conditions involving judgments of
positivity and counting syllables). Again, as in the Craik
et al. experiment, it was important to show that the various
tasks were comparable in terms of difficulty (as indexed by
response latency).
Ochsner, Beer, and their colleagues (full
disclosure: I was one of them!) reported just such a study in
2005, in which self-referent judgments were compared with
judgments involving the subjects' best friends (there
were also two control conditions involving judgments of
positivity and counting syllables). Again, as in the Craik
et al. experiment, it was important to show that the various
tasks were comparable in terms of difficulty (as indexed by
response latency).
 More
More  important, employing the same statistical
parametric mapping technique as Craik et al., Ochsner and Beer
found no brain area that was significantly more
activated by the self-referent processing task than by the
other-referent task. Self-reference produced more
activation in the medial prefrontal cortex compared to
syllable-counting, but other-reference produced the same
effect.
important, employing the same statistical
parametric mapping technique as Craik et al., Ochsner and Beer
found no brain area that was significantly more
activated by the self-referent processing task than by the
other-referent task. Self-reference produced more
activation in the medial prefrontal cortex compared to
syllable-counting, but other-reference produced the same
effect.
Of course, the logic of these studies depends on the assumption that the SRE is about self-reference. Unfortunately, research by Klein and his colleagues, summarized in the lecture supplement on The Self, indicates that the SRE is completely confounded with organizational activity. Put bluntly, it appears that the SRE has nothing to do with self-reference after all.
But I digress. In fact, between the Craik and the Ochsner/Beer studies a rather large literature had accumulated on the neural substrates of self-reference. Gillham and Farah (2005) reviewed a large number of studies involving judgments about the physical self (face recognition, body recognition, agency) and the psychological self (trait judgments, autobiographical memory retrieval, and taking a first-person perspective). In no instance did self-reference activate a different brain area, compared to other-reference.
 |
 |
 |
Perhaps the self is "just another person" after all!
 The specialization of function extends to the
two cerebral hemispheres, right and left, created by the central
commissure. Anatomically, the two hemispheres are
not quite identical: on average, the left hemisphere is slightly
larger than the right hemisphere. Most psychological
functions are performed by both hemispheres, but there is some
degree of hemispheric specialization as well. But in most
individuals, Broca's and Wernicke's areas, the parts of the
brain most intimately involved in language function, tend to be
localized in the left hemisphere -- though in some people the
right hemisphere does have some capacity for language as well.
The specialization of function extends to the
two cerebral hemispheres, right and left, created by the central
commissure. Anatomically, the two hemispheres are
not quite identical: on average, the left hemisphere is slightly
larger than the right hemisphere. Most psychological
functions are performed by both hemispheres, but there is some
degree of hemispheric specialization as well. But in most
individuals, Broca's and Wernicke's areas, the parts of the
brain most intimately involved in language function, tend to be
localized in the left hemisphere -- though in some people the
right hemisphere does have some capacity for language as well.
An interesting aspect of hemispheric specialization is contralateral projection, meaning that each hemisphere controls the functions of the opposite side of the body. Thus, the right hemisphere mediates sensorimotor functions on the left part of the body, auditory function of the left ear, and visual function of the left half-field of each eye. The left hemisphere does just the opposite. Ordinarily, the two hemispheres communicate with each other, and integrate their functions by means of the corpus callosum, a bundle of nerve fibers connecting them. But in cases of cerebral commisurotomy (a surgical procedure in which the corpus callosum has been severed), this communication is precluded. Remarkably enough, this situation rarely creates a problem: by virtue of eye movements, both hemispheres have access to the contents of both visual fields. So, the right hemisphere can "see" what's in the left visual field, and the left hemisphere can "see" what's in the right visual field.
However, if a stimulus is presented to the left (or right) stimulus field so briefly as to prevent it from being picked up by an eye movement, we can confine its processing to the contralateral hemisphere. This creates a situation where, quite literally, the left hemisphere does not know what the right hemisphere is doing, and vice-versa.
Michael Gazzaniga and his
colleagues have explored this situation by presenting different
pictures to each hemisphere, and then asking split-brain
patients to point to an associated picture in a larger array of
choices.
On the basis of results such as these, Gazzaniga has proposed that the left hemisphere is the site of a brain interpreter that is specialized for seeking explanations for internal and external events as a basis for making appropriate responses to them. In Gazzaniga's view, this brain module detects the relations between contiguous events, generates an explanation for the relationship, and then weaves that explanation into a "personal story".
In other words, Gazzaniga is proposing that certain structures in the left hemisphere are specialized for causal attribution -- one of the fundamental tasks of social cognition.
However, there is a problem with this proposal, which is that the left hemisphere also controls language function. It's possible that the right hemisphere also has the capacity to generate causal explanations, but simply can't express them through language.
Another area of social cognition
that has attracted neuropsychological interest is face
perception. Obviously, the face is a critical social
stimulus.
 Bruce, Young, and their colleagues have
offered an influential model of face perception involving a
number of different cognitive modules or systems:
Bruce, Young, and their colleagues have
offered an influential model of face perception involving a
number of different cognitive modules or systems:
Of particular interest to psychologists who study object perception is a neuropsychological syndrome known as visual object agnosia, in which patients can describe objects but cannot name them, recognize them as familiar, or demonstrate how they are used. A particular form of visual object agnosia is prosopagnosia, in which the patient can perceive and describe a face, but cannot recognize the face as familiar or name the person to whom the face belongs. Difficulties in face perception displayed by neurological patients were first described in the 19th century, by Charcot (1883) and Wilbrand (1892); a specific agnosia for faces was first described, and given the name prosopagnosia, by Bodamer (1947).
In terms of the Bruce & Young model of face perception, prosopagnosics appear to have impaired modules for face recognition and name generation (at least), but the module for structural description is spared.
 In the "pure" form of prosopagnosia, the
patient has difficulty recognizing familiar faces, but has no
difficulty with other objects. This suggests that there is
a brain system (or set of systems) that is specialized for the
perception of faces as opposed to other objects. In fact,
prosopagnosic patients typically show bilateral damage in a
portion of the visual association cortex at the boundary of the
occipital and temporal lobes (Brodmann's areas 18, 19, and
37). Kanwisher has named this area of the brain, centered
on the fusiform gyrus, the fusiform face area
(FFA).
In the "pure" form of prosopagnosia, the
patient has difficulty recognizing familiar faces, but has no
difficulty with other objects. This suggests that there is
a brain system (or set of systems) that is specialized for the
perception of faces as opposed to other objects. In fact,
prosopagnosic patients typically show bilateral damage in a
portion of the visual association cortex at the boundary of the
occipital and temporal lobes (Brodmann's areas 18, 19, and
37). Kanwisher has named this area of the brain, centered
on the fusiform gyrus, the fusiform face area
(FFA).
Actually, the identification of a specific area of the brain specialized for the perception of faces began with studies of inferotemporal (IT) cortex in monkeys. In the early 1960s, Charles Gross and his colleagues, employing the technique of single-unit recording, discovered that IT played a role in vision -- that is, certain neurons on IT fired when specific visual stimuli were presented to the animals (they also discovered analogous neurons in the superior temporal cortex responsive to auditory stimulation). As research progressed, they stumbled on a bundle of neurons that failed to respond to any of the visual stimuli presented to them so far. More or less by accident, they waved their hands in front of the screen -- and the neurons responded. In 1969, Gross and his colleagues reported that these cells were specifically responsive to hands -- in particular, monkey hands. In 1984, Gross and his colleagues reported another bundle of IT neurons that responded specifically to faces -- and, in particular, to monkey faces. As Gross recounts the story, this research went virtually unnoticed by the wider field of neurophysiology for more than a decade ("Processing the Facial Image: A Brief History" by C.G. Gross, American Psychologist, 60, 755-763, 2005).
 Based on neuropsychological
studies of prosopagnosic patients, Justine Sergent (1992) had
identified the fusiform area as critical for face
perception. Her ideas were confirmed by Kanwisher's
neuroimaging studies of neurologically intact subjects.
Based on neuropsychological
studies of prosopagnosic patients, Justine Sergent (1992) had
identified the fusiform area as critical for face
perception. Her ideas were confirmed by Kanwisher's
neuroimaging studies of neurologically intact subjects.
 Given the status of humans as
social animals, and the critical importance of the face for
social behavior, it makes sense that a specific area of the
brain might have evolved to process information about the face
-- just as other areas of the brain, such as Broca's and
Wernicke's areas, have evolved to process information about
speech and language. And precisely because the proposal
makes evolutionary sense, it has been widely accepted. As
Kanwisher put it: Face Recognition is "a cognitive function with
its own private piece of real estate in the brain" -- namely,
the fusiform gyrus.
Given the status of humans as
social animals, and the critical importance of the face for
social behavior, it makes sense that a specific area of the
brain might have evolved to process information about the face
-- just as other areas of the brain, such as Broca's and
Wernicke's areas, have evolved to process information about
speech and language. And precisely because the proposal
makes evolutionary sense, it has been widely accepted. As
Kanwisher put it: Face Recognition is "a cognitive function with
its own private piece of real estate in the brain" -- namely,
the fusiform gyrus.
Further analysis has suggested that the FFA
consists of at least six separate areas, or "face patches",
each activated in response to a particular feature of the
face, such as the separation between the eyes.
In the first place, prosopagnosia
is not always, or even often, specific to faces. Most
prosopagnosics have deficits in recognizing objects other than
faces, too.
 Another,
more important problem is that the standard tests of prosopagnosia
typically contain a confound.
Another,
more important problem is that the standard tests of prosopagnosia
typically contain a confound. A
better comparison would be to ask patients not only to recognize
particular faces, but also to recognize particular objects --
their own house as opposed to a neighbor's, or the White House
as opposed to Buckingham Palace, for example. This
rigorous comparison, which controls for the level of object
categorization, is rarely performed.
A
better comparison would be to ask patients not only to recognize
particular faces, but also to recognize particular objects --
their own house as opposed to a neighbor's, or the White House
as opposed to Buckingham Palace, for example. This
rigorous comparison, which controls for the level of object
categorization, is rarely performed.
Coupled with her argument about level of categorization is an argument about expertise. Novices in a domain tend to identify objects in that domain at the basic-object level of categorization -- to say "bird" when we see a bird, "cat" when we see a cat. But bird and cat experts don't do this. They identify objects at subordinate levels of classification -- they say "robin" or "sparrow", or "calico" or "Blue-point Siamese".
 The acquisition of
expertise in a domain is indicated by the entry-level shift:
At initial stages of training, novices are much slower to
classify objects at the subordinate level, compared to the basic
level. But as training proceeds, classification speed
progressively diminishes, so that experts classify objects at
the subordinate level as quickly as they classify them at the
basic level. The entry-level shift, therefore, provides an
objective index of expertise.
The acquisition of
expertise in a domain is indicated by the entry-level shift:
At initial stages of training, novices are much slower to
classify objects at the subordinate level, compared to the basic
level. But as training proceeds, classification speed
progressively diminishes, so that experts classify objects at
the subordinate level as quickly as they classify them at the
basic level. The entry-level shift, therefore, provides an
objective index of expertise.
In fact, Isabel Gauthier, Michael Tarr, and their colleagues (Gauthier was Tarr's graduate student at Yale; she is now at Vanderbilt, he is now at Carnegie-Mellon) have performed a number of interesting studies, employing both experimental and brain-imaging methods with both normal subjects and brain-damaged patients, which strongly indicate that the fusiform area is not dedicated to face recognition, but rather is specialized for classification at subordinate levels of categorization. more general deficit in recognizing objects at a particular (subordinate) level of categorization.
 Some
Some  of these
experiments involved "natural" experts in cars and birds.
Consider, for example, a brain-imaging study of visual object
classification, in which subjects were asked to classify objects
at various levels of categorization (Gauthier, Anderson, Tarr,
& Sudlarski, 1997). For example, they might be
presented with a picture of a bird and then asked one of two
questions: Is it a bird? -- a basic-object-level
question; and Is it a pelican? -- a
subordinate-level question. The basic finding of the
experiment is that classifying birds at subordinate-level of
categorization activated the same "fusiform face area" that is
involved in face recognition.
of these
experiments involved "natural" experts in cars and birds.
Consider, for example, a brain-imaging study of visual object
classification, in which subjects were asked to classify objects
at various levels of categorization (Gauthier, Anderson, Tarr,
& Sudlarski, 1997). For example, they might be
presented with a picture of a bird and then asked one of two
questions: Is it a bird? -- a basic-object-level
question; and Is it a pelican? -- a
subordinate-level question. The basic finding of the
experiment is that classifying birds at subordinate-level of
categorization activated the same "fusiform face area" that is
involved in face recognition.
 In
another
experiment, bird experts activated the fusiform area when they
classified birds but not cars, while car experts activated the
fusiform area when they classified cars but not birds.
In
another
experiment, bird experts activated the fusiform area when they
classified birds but not cars, while car experts activated the
fusiform area when they classified cars but not birds.
 In
In  other studies, subjects learned to classify
artificial objects, designed by Scott Yu, working in Tarr's
laboratory, known as "greebles" (a name coined by Robert
Abelson, a distinguished social psychologist who was Tarr's
colleague at Yale). The greebles are designed so that they
fall into several "families", defined by body shape, and two
"genders", defined by whether their protuberances pointed up or
down. Greeble novices activated the FFA when they
identified faces, but not greebles. But when they became
greeble experts, the same subjects activated the FFA when they
classified both kinds of objects.
other studies, subjects learned to classify
artificial objects, designed by Scott Yu, working in Tarr's
laboratory, known as "greebles" (a name coined by Robert
Abelson, a distinguished social psychologist who was Tarr's
colleague at Yale). The greebles are designed so that they
fall into several "families", defined by body shape, and two
"genders", defined by whether their protuberances pointed up or
down. Greeble novices activated the FFA when they
identified faces, but not greebles. But when they became
greeble experts, the same subjects activated the FFA when they
classified both kinds of objects.
Lest anyone think that greebles are "face-like", and thus that the comparison is unfair, Gauthier and her colleagues have shown that a particular neurological patient, M.K., who suffers from a visual object agnosia that nonetheless spares face recognition, cannot classify greebles. Apparently, from the point of view of M.K.'s brain, greebles are not face-like stimuli.
 In other studies, the subjects learned to
classify snowflakes, and fingerprints, with the same
results.
In other studies, the subjects learned to
classify snowflakes, and fingerprints, with the same
results.
| Yes, every snowflake is different, just as fingerprints are, but there are ways to classify snowflakes based on shared properties. |  |
| And since you asked: yes, there are ways of classifying fingerprints, too. |  |
The general finding of these studies is that subordinate-level classification activated the fusiform area, whether the task involved recognizing faces, greebles, or snowflakes (or birds or automobiles). Moreover, fusiform activation increased as subjects became "experts" at making the classification judgments required of them.
 On the basis of these sorts of
findings, Gauthier and Tarr have proposed that the area
surrounding the fusiform gyrus be considered to be a flexible
fusiform area (not coincidentally, also abbreviated FFA),
which is involved in classification of all sorts of objects at
the subordinate level of categorization. Faces are one
example of such stimuli, but there are others. The point
is that while face recognition epitomizes subordinate-level
classification, other recognition tasks can involve
subordinate-level classification as well.
On the basis of these sorts of
findings, Gauthier and Tarr have proposed that the area
surrounding the fusiform gyrus be considered to be a flexible
fusiform area (not coincidentally, also abbreviated FFA),
which is involved in classification of all sorts of objects at
the subordinate level of categorization. Faces are one
example of such stimuli, but there are others. The point
is that while face recognition epitomizes subordinate-level
classification, other recognition tasks can involve
subordinate-level classification as well.
Gauthier and Tarr's challenge to the traditional interpretation of the Fusiform Face Area has not gone unchallenged. One serious problem is that of spatial blurring. That is, the relatively low spatial resolution of standard fMRI (SR-fMRI), typically used in the expertise studies, may not be able to distinguish that portion of the fusiform "real estate" from another area, actually a separate brain module, that performs expert classification in other domains. Put another way, the non-face-selective regions may border the true FFA, and very high-resolution brain imaging may be needed to separate them.
Now, this looks like a reach, but in fact the expertise studies have used SR-fMRI, which has a relatively low spatial resolution. And there are several studies using high-resolution fMRI (HR-fMRI), which documenting an FFA which doesn't seem to respond to non-faces. At the same time, these HR-fMRI studies haven't taken account of expertise. All they've done is to ask ordinary people to classify cars or birds. So you can see where this is going. What's needed is an HR-fMRI study that takes account of expertise in the non-face domain.
 Such
a study was recently performed
by Randal McGugin, working in
Gauthier's laboratory (McGugin
et al., 2012). They
employed a 7-Tesla (7T) MRI scanner,
pretty much the most powerful
scanner currently available,
and ran it at both SR-fMRI and
HR-fMRI settings.
Their subjects represented
varying degrees of expertise
at automobile
identification, as
assessed behaviorally
prior to the
scanning runs
(remember,
everyone's an
expert at face-identification).
During the
scanning runs
themselves, they
were presented
with five
classes of stimuli
-- faces, animals,
automobiles, airplanes,
and scrambled images
(these last
served as
controls).
Two images
were presented
at once, and
they were
asked to
determine
whether they
belonged to
the same
person, same
make
and model of
car,
etc.
Such
a study was recently performed
by Randal McGugin, working in
Gauthier's laboratory (McGugin
et al., 2012). They
employed a 7-Tesla (7T) MRI scanner,
pretty much the most powerful
scanner currently available,
and ran it at both SR-fMRI and
HR-fMRI settings.
Their subjects represented
varying degrees of expertise
at automobile
identification, as
assessed behaviorally
prior to the
scanning runs
(remember,
everyone's an
expert at face-identification).
During the
scanning runs
themselves, they
were presented
with five
classes of stimuli
-- faces, animals,
automobiles, airplanes,
and scrambled images
(these last
served as
controls).
Two images
were presented
at once, and
they were
asked to
determine
whether they
belonged to
the same
person, same
make
and model of
car,
etc.
 |
 |
 |
 |
The results of the study are a little complicated, but here's the gist of it:
All of which seems to
suggest that Tarr and Gauthier had it pretty much right the
first time, and that the "FFA" isn't really selective for faces,
but rather is specialized for subordinate-level
object-identification, of which face recognition is the best and
most familiar example. Still, McGugin's experiment
illustrates the complexity of the project of doing cognitive
neuroscience right. If it's this hard to identify a
specific brain module for something as plausibly innate as
ready-for-modularity as face recognition, think how hard it's
going to be for other social-cognitive tasks!
Gauthier and Tarr's reconstrual of the FFA remains somewhat controversial, and further research may lead them to revisit their conclusions. Still, Gauthier's work is good enough that in 2008 she received the Troland Award from the National Academy of Sciences, which honors outstanding young scientists in the field of psychology.
 Some evidence in
this regard comes from a new study by Quian Quiroga et al. (Neuron, 2014), employing the
same single-unit recording methodology employed in their study of "grandmother neurons",
described in the lectures on "Social
Memory" (see also "Brain
Cells for Grandmother" by
Rodrigo Quian Quiroga, Itzhak Fried, & Christoph
Koch, Scientific American, 02/2013).
The research involved patients undergoing surgical
treatment for epilepsy, who had micro-electrodes
embedded in various portions of their temporal cortex
in order to identify the point of origin of their
seizures. As in the earlier study, Quian Quiroga et al. presented these
patients with pictures of familiar people (e.g.,
Jennifer Aniston) and locations (e.g., the
Sydney Opera House) and identified a number of
locations that responded
specifically to particular
stimuli. For example, they
identified in one patient a unit that responded
selectively to photographs of
President Bill Clinton, but not to photos of George
Bush. They then created a
composite of the two presidents' faces (call it
"Clintush"). As the image switched from Clinton
to Bush, the unit ceased activity. They then looked at responses
to the "Clintush" composite. When
subjects are first presented
with a picture of Clinton, they perceived
"Clintush" as Bush, and vice-versa -- a
phenomenon of sensory adaptation. The
"Clinton neuron" responded
accordingly. When the subject
perceived the composite as Clinton, the unit became active; when he
perceived it as Bush, it was
inactive. While there are
regions of fusiform cortex that are
selectively responsive to particular
faces, as Kanwisher argues, other
fusiform areas are responsive to other
categories of stimuli, as Gauthier argues.
But there also appear to be individual eurons (or,
more likely, small clusters of neurons) in the fusiform cortex that are
differentially responsive to particular faces. (Image
taken from "The Face as Entryway to the Self" by
Christof Koch, one of the co-authors of the
Quian-Quiroga studies, Scientific
American Mind, 01-02/2014.)
Some evidence in
this regard comes from a new study by Quian Quiroga et al. (Neuron, 2014), employing the
same single-unit recording methodology employed in their study of "grandmother neurons",
described in the lectures on "Social
Memory" (see also "Brain
Cells for Grandmother" by
Rodrigo Quian Quiroga, Itzhak Fried, & Christoph
Koch, Scientific American, 02/2013).
The research involved patients undergoing surgical
treatment for epilepsy, who had micro-electrodes
embedded in various portions of their temporal cortex
in order to identify the point of origin of their
seizures. As in the earlier study, Quian Quiroga et al. presented these
patients with pictures of familiar people (e.g.,
Jennifer Aniston) and locations (e.g., the
Sydney Opera House) and identified a number of
locations that responded
specifically to particular
stimuli. For example, they
identified in one patient a unit that responded
selectively to photographs of
President Bill Clinton, but not to photos of George
Bush. They then created a
composite of the two presidents' faces (call it
"Clintush"). As the image switched from Clinton
to Bush, the unit ceased activity. They then looked at responses
to the "Clintush" composite. When
subjects are first presented
with a picture of Clinton, they perceived
"Clintush" as Bush, and vice-versa -- a
phenomenon of sensory adaptation. The
"Clinton neuron" responded
accordingly. When the subject
perceived the composite as Clinton, the unit became active; when he
perceived it as Bush, it was
inactive. While there are
regions of fusiform cortex that are
selectively responsive to particular
faces, as Kanwisher argues, other
fusiform areas are responsive to other
categories of stimuli, as Gauthier argues.
But there also appear to be individual eurons (or,
more likely, small clusters of neurons) in the fusiform cortex that are
differentially responsive to particular faces. (Image
taken from "The Face as Entryway to the Self" by
Christof Koch, one of the co-authors of the
Quian-Quiroga studies, Scientific
American Mind, 01-02/2014.)
Perhaps definitive
evidence comes from research on the perception of
(human) faces in macaques by Le Chang and Doris
Tsao at Cal Tech (Cell, 2017;
see also the commentary by Quian Quiroga
accompanying the article; for a less technical
account of this research, see "Face Values" by
Doris Tsao, Scientific American, 02/2019).
Employing a variant
of Hubel and Wiesel's classic technique, they
recorded from single units in the "face patches"
of the fusiform area -- areas of adjacent neurons (there are maybe
six of them) that appear to be
particularly responsive to faces. Instead of
presenting individual faces (familiar or not), as
Quian Quiroga did in the "Jennifer Anniston"
study, Chang and Tsao systematically varied
various features of the faces, such as roundness,
interocular distance, hairline width, skin tone,
and skin texture. They found that individual
cells responded to particular features -- or, more
precisely, to higher-order combinations of
particular features. C&T argue that
individual faces can be located at points in a
multidimensional space (consisting of 50 or more
dimensions!), and individual neurons respond to
departures from the average value on each of these
dimensions. If a face does not present the
particular combination of features to which an
individual cell has been "tuned", it will not
respond to it; but it will respond to different
faces that have that particular higher-order
feature. Chang and Tsao estimate that about
200 individual neurons are required to uniquely
identify each face -- a different combination of
neurons for each face. Then, employing
techniques pioneered by UCB's Jack Gallant, they
were able to reconstruct individual faces from
recordings of the activity of a population of
about 200 neurons -- not just the front view, but
also the profile view as well. It's a great
paper; even the Scientific American
version is a great read.
So here's the
situation as it now stands (I think). There
isn't a "fusiform face area", exactly, though
there are anatomically segregated patches of
neurons that are particularly sensitive to
faces. Individual neurons within each of
these patches respond to variations in facial
features along 50 or more dimensions. The
particular pattern with which these neurons fire
is the neural signature of an individual
face. In addition, faces that are very
familiar are also represented by individual
neurons, or perhaps small groups of adjacent
neurons, in the hippocampus -- much the way other
memories might be.
 In a series of
studies, a group of Italian neuroscientists studied the behavior
of individual neurons while macaque monkeys engaged in various
activities, such as grasping a piece of fruit. The
initial goal of this research was to extend the line of research
initiated by Barlow, Lettvin, and Hubel and Wiesel (discussed in
the lectures on "Social
Memory" from the perceptual domain to the domain of
action. Recall that Hubel and Wiesel discovered single
cells in the frog's visual system that appeared to respond to
particular features of the stimulus -- edges, points of light
and dark, and the like -- "bug detectors", if you will.
Giacomo Rizzolatti and his colleagues (1988) at the University
of Parma, in Italy, were doing much the same thing with
action. Recording from single cells in the ventral
premotor cortex in macaque monkeys, they tried to identify
particular neurons that fired whenever the animals performed
specific actions with their hands or mouths -- reaching for an
object, for example, or grasping or manipulating it. In
theory, activation of these clusters of neurons enable the
organism to perform the specified action automatically.
In a series of
studies, a group of Italian neuroscientists studied the behavior
of individual neurons while macaque monkeys engaged in various
activities, such as grasping a piece of fruit. The
initial goal of this research was to extend the line of research
initiated by Barlow, Lettvin, and Hubel and Wiesel (discussed in
the lectures on "Social
Memory" from the perceptual domain to the domain of
action. Recall that Hubel and Wiesel discovered single
cells in the frog's visual system that appeared to respond to
particular features of the stimulus -- edges, points of light
and dark, and the like -- "bug detectors", if you will.
Giacomo Rizzolatti and his colleagues (1988) at the University
of Parma, in Italy, were doing much the same thing with
action. Recording from single cells in the ventral
premotor cortex in macaque monkeys, they tried to identify
particular neurons that fired whenever the animals performed
specific actions with their hands or mouths -- reaching for an
object, for example, or grasping or manipulating it. In
theory, activation of these clusters of neurons enable the
organism to perform the specified action automatically.
Subsequently, these investigators discovered that these same neurons also fired when the monkeys, albeit restrained, merely observed such actions displayed by another organism (including, according to a famous "origin myth", a human research assistant licking a cone of gelato). DePelligrino et al. (1992) and Gallese et al. (1996) dubbed these cells mirror neurons, because it seemed that the neurons were reflecting actions performed by other organisms.
 Further
search located another set of mirror neurons in the inferior
portion of the parietal lobe -- an area that is connected to the
premotor cortex. This makes sense, because while the
frontal lobe controls motor activity (such as reaching) by
various parts of the body, the parietal lobe processes
somatosensory information from those same body parts.
Further
search located another set of mirror neurons in the inferior
portion of the parietal lobe -- an area that is connected to the
premotor cortex. This makes sense, because while the
frontal lobe controls motor activity (such as reaching) by
various parts of the body, the parietal lobe processes
somatosensory information from those same body parts.
Based largely
on a principle of evolutionary continuity,
it seems reasonable to expect that humans have mirror
neurons, too. However, practical and ethical
considerations make it difficult (though not impossible) to
perform single-unit recordings of human brain
activity. Instead,
researchers have used various
brain-imaging techniques to look for activations in
the frontal and parietal lobes when
humans observe other humans in action. Because we only know what is
happening to larger brain regions, as opposed to
individual neurons, these
regions are generally considered to be mirror
neuron "systems" (MNSs).
 Regions of the
frontal and parietal lobes have been designated as a
mirror neuron system for action.
Regions of the
frontal and parietal lobes have been designated as a
mirror neuron system for action.
 There also
appears to be a mirror neuron system for emotion,
revealed when subjects observe the emotional
reactions of another
person, such as pain or disgust.
There also
appears to be a mirror neuron system for emotion,
revealed when subjects observe the emotional
reactions of another
person, such as pain or disgust.
 Here's an elegant
study that reveals the operation of the mirror
neuron system for action in humans. Calvo-Merino
et al. (2004) used fMRI to record brain activity in skilled
dancers while they watched other dancers perform. Some
of the subjects were professional ballet
dancers; others were practitioners of capoeira, a
Brazilian martial art that has some dance elements to it;
there were also control subjects who had no skill in either dance form. The
frontal and parietal MNSs was activated when ballet
dancers watched other ballet dancers, but not when they
watched capoeira dancers; the MNSs of the capoeira
dancers were activated when they watched
other capoeira dancers, but not when they watched
ballet dancers. In other words, the MNSs were
activated only when the subjects watched other people
perform actions which they also knew how to perform.
Here's an elegant
study that reveals the operation of the mirror
neuron system for action in humans. Calvo-Merino
et al. (2004) used fMRI to record brain activity in skilled
dancers while they watched other dancers perform. Some
of the subjects were professional ballet
dancers; others were practitioners of capoeira, a
Brazilian martial art that has some dance elements to it;
there were also control subjects who had no skill in either dance form. The
frontal and parietal MNSs was activated when ballet
dancers watched other ballet dancers, but not when they
watched capoeira dancers; the MNSs of the capoeira
dancers were activated when they watched
other capoeira dancers, but not when they watched
ballet dancers. In other words, the MNSs were
activated only when the subjects watched other people
perform actions which they also knew how to perform.
The discovery of mirror neurons has important implications for theories of the neural substrates of visual perception. Ordinarily, we would think that the recognition of visible actions would be accomplished by the occipital lobe, which is the primary visual cortex. But it appears that the same neural system that is involved in generating actions -- the premotor cortex -- is also involved in recognizing them. At least so far as perceiving actions is concerned, there appears to be more to perception than vision. Perception of action also recruits parts of the brain that are critical for producing action. Put more formally, it appears that perception and action share common representational formats in the brain. The same areas of the brain are involved in perceiving bodily movements as in producing them.
At the same time, it has been suggested that mirror neurons may be important for monkeys' ability to imitate and understand each other's behavior. If the same neurons fire when the monkey grasps a cup as when a monkey sees another monkey grasp a cup, the first monkey can understand the second monkey's behavior by saying to itself, as it were, "this is what I'd be doing if I did that".
Mirror neurons have also been discovered in the perception of facial actions, suggesting that they may play a role in the perception of facial expressions of emotion -- "this is what I'd be feeling if I looked like that".
Which is a very informal way of illustrating the ideomotor framework for action, which holds that perceiving an action and performing an action are closely linked.
A human example of the ideomotor framework for action is Alvin Liberman's motor theory of speech perception, which argues that, instead of identifying patterns of speech sounds as such, the listener identifies the actions in the motor tract -- movements of the lips, tongue, larynx, and the like -- that would produce those sounds -- "this is what I would be saying if I did that".
 It did not take long
for various theorists to suggest that mirror neurons played a
critical role in human social cognition -- in particular, our
ability to understand and to empathize with each other's
emotional states. Unfortunately, documenting mirror
neurons in humans is made difficult by the virtual impossibility
of doing single-unit recording in living human brains (yes,
Quian-Quiroga did it, but only in the special circumstance that
their subjects were undergoing necessary brain surgery
anyway). Instead, researchers have had to be content with
neuroimaging studies of brain activity while subjects are
engaged, like the monkeys, in observing other people's actions
and facial expressions. Neuroimaging does not have the
spatial resolution that would be required to identify particular
neurons associated with particular actions, but they can
identify the general locations in the brain where such neurons
would like. And, in fact, Iacoboni et al. (1999)
identified homologous structures in the human brain -- in the
inferior frontal gyrus and the inferior parietal lobule -- that
are active when observing other people's actions or facial
expressions. These areas of the brain are now called the
frontal and parietal mirror neuron systems.
It did not take long
for various theorists to suggest that mirror neurons played a
critical role in human social cognition -- in particular, our
ability to understand and to empathize with each other's
emotional states. Unfortunately, documenting mirror
neurons in humans is made difficult by the virtual impossibility
of doing single-unit recording in living human brains (yes,
Quian-Quiroga did it, but only in the special circumstance that
their subjects were undergoing necessary brain surgery
anyway). Instead, researchers have had to be content with
neuroimaging studies of brain activity while subjects are
engaged, like the monkeys, in observing other people's actions
and facial expressions. Neuroimaging does not have the
spatial resolution that would be required to identify particular
neurons associated with particular actions, but they can
identify the general locations in the brain where such neurons
would like. And, in fact, Iacoboni et al. (1999)
identified homologous structures in the human brain -- in the
inferior frontal gyrus and the inferior parietal lobule -- that
are active when observing other people's actions or facial
expressions. These areas of the brain are now called the
frontal and parietal mirror neuron systems.
 Mirror neurons are quickly
becoming the hottest topic in social neuroscience. V.S. Ramachandran, one of the world's
leading cognitive neuroscientists, have called mirror neurons
"the driving force behind the 'great leap forward' in human
evolution" -- that is, they are the
neurological difference between humans, and our closest
evolutionary relatives, and the rest of the
animal world.
Mirror neurons are quickly
becoming the hottest topic in social neuroscience. V.S. Ramachandran, one of the world's
leading cognitive neuroscientists, have called mirror neurons
"the driving force behind the 'great leap forward' in human
evolution" -- that is, they are the
neurological difference between humans, and our closest
evolutionary relatives, and the rest of the
animal world.
 As
noted, they've been implicated in the perception of facial
expressions of emotion, and also in empathy. They've been
implicated in imitation, which is a basic form of social
learning (see the lectures on "Personality and
Social Cognition"). It's been suggested that they
comprise the neural substrate of the "theory of mind" by
which we understand other people's beliefs, desires, and
feelings (see the lectures on "Social-Cognitive
Development"). It's been suggested (by V.S.
Ramachandran) that autistic individuals, who seem to have
specific deficits in social cognition, have suffered insult,
injury, or disease affecting their motor neuron systems (for
more detail, see the lectures on "Social-Cognitive
Development"). And, of course, it's been suggested
that girls and women have more, or more developed, mirror
neurons than boys and men. Greg Hickok has called mirror
neurons "the rock stars of cognitive neuroscience".
As
noted, they've been implicated in the perception of facial
expressions of emotion, and also in empathy. They've been
implicated in imitation, which is a basic form of social
learning (see the lectures on "Personality and
Social Cognition"). It's been suggested that they
comprise the neural substrate of the "theory of mind" by
which we understand other people's beliefs, desires, and
feelings (see the lectures on "Social-Cognitive
Development"). It's been suggested (by V.S.
Ramachandran) that autistic individuals, who seem to have
specific deficits in social cognition, have suffered insult,
injury, or disease affecting their motor neuron systems (for
more detail, see the lectures on "Social-Cognitive
Development"). And, of course, it's been suggested
that girls and women have more, or more developed, mirror
neurons than boys and men. Greg Hickok has called mirror
neurons "the rock stars of cognitive neuroscience".
 Based on studies
of mirror neurons and mirror neuron systems, Gallese and his
colleagues (2004) have proposed a unifying neural
hypothesis of social cognition. That is,
mirror neurons are the biological
substrate of social cognition, creating "a bridge... between others and ourselves".
Based on studies
of mirror neurons and mirror neuron systems, Gallese and his
colleagues (2004) have proposed a unifying neural
hypothesis of social cognition. That is,
mirror neurons are the biological
substrate of social cognition, creating "a bridge... between others and ourselves".
In this way, Gallese et al. argue, the
activity of the mirror neuron system doesn't
simply support thinking about other people (though it
does that too); it leads to experiential
insight into them.
 Maybe. Frankly, it's just
too early to tell. As Alison Gopnik has noted ("Cells That
Read Minds? in Slate, 04/26/2007), the enthusiasm for
mirror neurons is reminiscent of another enthusiasm, that marked
the very beginnings of cognitive neuroscience -- the distinction
between the "right brain" and the "left brain". Sure,
there is considerable hemispheric specialization -- not least,
language function appears to be lateralized to the left cerebral
hemisphere. But that doesn't warrant speculation that the
right hemisphere is the seat of the unconscious, or that men,
with their alleged propensities for "linear thinking" are
"left-brained", while women, with their holistic tendencies, are
"right-brained".
Maybe. Frankly, it's just
too early to tell. As Alison Gopnik has noted ("Cells That
Read Minds? in Slate, 04/26/2007), the enthusiasm for
mirror neurons is reminiscent of another enthusiasm, that marked
the very beginnings of cognitive neuroscience -- the distinction
between the "right brain" and the "left brain". Sure,
there is considerable hemispheric specialization -- not least,
language function appears to be lateralized to the left cerebral
hemisphere. But that doesn't warrant speculation that the
right hemisphere is the seat of the unconscious, or that men,
with their alleged propensities for "linear thinking" are
"left-brained", while women, with their holistic tendencies, are
"right-brained".
It's possible that mirror neurons
are responsible for everything that makes us human. But
then again...
Besides, it's not even clear that
mirror neurons are critical for action perception in macaques (see
G. Hickok, "Eight Problems for the Mirror Neuron Theory of Action
Understanding in Monkeys and Humans, Journal of Cognitive
Neuroscience, 2009).
The critical point about mirror
neurons is that they are critical for understanding action.
But Hickok (2011) argues that mirror neurons don't have the
semantic that would be required for understanding
action. Those semantics are stored elsewhere in the brain --
along with everything else that we know. Hickok points out
that we can understand actions without being able to execute them,
as in the example of Broca's aphasia above. More
prosaically, he points out that we can understand spectator sports
without ourselves being able to perform the motor actions we
observe the players enacting. As an alternative, Hickok
argues that the MNS plays a role in action selection,
supporting sensorimotor integration -- that is, with integrating
what we observe with what we do in response. In his view,
the MNS is activated not just by any observation, but rather by
observation in conjunction with the observer's own goals.
The MNS becomes involved only because the actions of others are
relevant for our selection of our own actions.
Do Mirror Neurons Make Us Human?In The Tell-Tale Brain: A Neuroscientist's Quest for What Makes us Human (2011), V.S. Ramachandran argues that mirror neurons will play the same role in psychology as the discovery of DNA plays in biology. Reviewing the book, however, the philosopher Anthony Gottleib offers a demurral ("A Lion in the Undergrowth", New York Times Book Review, 01/30/2011):
|
 Similar
considerations apply to another class of neurons that has been
implicated in social cognition, known as von Economo
neurons (VENs) after Constantin von Economo, an Austrian
neuro-psychiatrist who first described them in 1925. In
the course of his cytoarchitectonic studies of brain tissue (a
failed attempt to supplant Brodmann's 1909 mapping of the
brain), von Economo identified a large neuron with an unusual
spindle shape, and a very sparse dendritic tree, located
exclusively in two areas of the frontal lobe: the anterior
cingulate cortex (ACC, Brodmann Area 24) and the
fronto-insular cortex (Brodmann's area 12). These are
areas of the brain that are damaged in fronto-temporal
dementia, a form of dementia. Unlike Alzheimer's
disease, in which the primary symptoms involve memory
functions, fronto-temporal dementia is characterized by marked
deficits in social and emotional functioning.
Similar
considerations apply to another class of neurons that has been
implicated in social cognition, known as von Economo
neurons (VENs) after Constantin von Economo, an Austrian
neuro-psychiatrist who first described them in 1925. In
the course of his cytoarchitectonic studies of brain tissue (a
failed attempt to supplant Brodmann's 1909 mapping of the
brain), von Economo identified a large neuron with an unusual
spindle shape, and a very sparse dendritic tree, located
exclusively in two areas of the frontal lobe: the anterior
cingulate cortex (ACC, Brodmann Area 24) and the
fronto-insular cortex (Brodmann's area 12). These are
areas of the brain that are damaged in fronto-temporal
dementia, a form of dementia. Unlike Alzheimer's
disease, in which the primary symptoms involve memory
functions, fronto-temporal dementia is characterized by marked
deficits in social and emotional functioning.
It was originally thought that VENs appeared uniquely in hominid brains (e.g., chimpanzees as opposed to macaques), but they have now been found in the brains of other social species, such as elephants. Still, the presence of VENs in the brains of social animals, especially in regions of the brain associated with social and emotional processes (such as the ACC and the frontal insula), has suggested that they are part of the neural substrate of social intelligence.
Maybe, but again, at least for now, this is little more than a good story.
In the first place, we don't really know what the functions of VENs are. For that matter, we don't know what the function of the ACC is -- does it monitor conflict, or correct errors, is it part of the reward system, or something else? The best guess about the ACC is that it's involved in everything that's interesting.
We don't even know if VENs are structurally different from other neurons. As Terry Deacon has noted, it is entirely possible that they're just on the "high" end in terms of size (also also more spindle-like, less pyramidal), and on the "low" end in terms of the number of dendrites. It's interesting that VENs are concentrated in the ACC, but that may have more to do with their physiological properties than with their psychological function: by virtue of their size, VENs conduct neural impulses very rapidly; and by virtue of their sparse dendritic foliage, they conduct neural impulses very selectively. Maybe these are good properties for neurons in a brain system that processes conflict.
 Just as social neuroscience began to settle into the
search for dedicated social-cognitive modules in the brain, a
new approach to brain imaging has begun to emerge, known as brain
mapping (Gallant et al., 2011). The essential
feature of brain mapping is that records the entire activity of
the brain as the subject performs some task -- not just a
relatively circumscribed region of interest, such as the
fusiform gyrus.
And then investigators
attempt to reconstruct the stimulus from the entire pattern of
brain activity. So far, brain-mapping has been attempted with various
perceptual tasks that are not particularly social in
nature. But as a matter of
historical fact, social neuroscience lags cognitive neuroscience
by about 5-10 years, so it is only a matter of time before
social neuroscientists turn to brain-mapping as well.
Just as social neuroscience began to settle into the
search for dedicated social-cognitive modules in the brain, a
new approach to brain imaging has begun to emerge, known as brain
mapping (Gallant et al., 2011). The essential
feature of brain mapping is that records the entire activity of
the brain as the subject performs some task -- not just a
relatively circumscribed region of interest, such as the
fusiform gyrus.
And then investigators
attempt to reconstruct the stimulus from the entire pattern of
brain activity. So far, brain-mapping has been attempted with various
perceptual tasks that are not particularly social in
nature. But as a matter of
historical fact, social neuroscience lags cognitive neuroscience
by about 5-10 years, so it is only a matter of time before
social neuroscientists turn to brain-mapping as well.
 The
emergence of neuropsychological and neuroscientific methods and
theories into the study of social cognition underscores the fact
that human experience, thought, and action can be analyzed at
three quite different levels:
The
emergence of neuropsychological and neuroscientific methods and
theories into the study of social cognition underscores the fact
that human experience, thought, and action can be analyzed at
three quite different levels:
Part of the appeal of cognitive
and social neuroscience is that it promises to connect
psychology "down" to the biophysical level of analysis, shedding
light on the neural substrates of cognition and social
interaction. But some neuroscientists have offered another
rationale, which is that neuroscientific evidence can affect
theorizing at the psychological and even sociocultural levels of
analysis. The general idea is that neuroscientific
findings will constrain psychological theories, forcing
us to choose, between competing theories, which one is right
based on its compatibility with neuroscientific evidence.
 The idea
of constraint was explicitly evoked by Gazzaniga in his
statement of the agenda for cognitive neuroscience.
Recall that Marr had proposed that the
implementational level could be attacked only after work had been completed at the computational
and algorithmic levels of analysis. But Gazzaniga
supposes that evidence of physical
implementation can shape theory at the computational and
algorithmic levels, as well.
The idea
of constraint was explicitly evoked by Gazzaniga in his
statement of the agenda for cognitive neuroscience.
Recall that Marr had proposed that the
implementational level could be attacked only after work had been completed at the computational
and algorithmic levels of analysis. But Gazzaniga
supposes that evidence of physical
implementation can shape theory at the computational and
algorithmic levels, as well.
 The
The
 rhetoric of constraint
comes up in an even earlier argument for cognitive
neuroscience, by Stephen Kosslyn, which drew
an analogy to architecture (but see if you can
spot the weakness of the analogy). A
little later, writing with Kevin Ochsner, he
invoked a version of Marr's
three-level organization of cognitive
research -- but, like
Gazzaniga, he argued that explanations at
the computational and algorithmic levels
depended on the details
at the
implementational level..
rhetoric of constraint
comes up in an even earlier argument for cognitive
neuroscience, by Stephen Kosslyn, which drew
an analogy to architecture (but see if you can
spot the weakness of the analogy). A
little later, writing with Kevin Ochsner, he
invoked a version of Marr's
three-level organization of cognitive
research -- but, like
Gazzaniga, he argued that explanations at
the computational and algorithmic levels
depended on the details
at the
implementational level..
This viewpoint
has sometimes been carried over into social-cognitive
neuroscience.
 For example, in an early paper,
Cacioppo and Berntsen (1992) asserted that "knowledge of the
body and brain can usefully constrain and inspire concepts and
theories of psychological function." In
a more recent advertisement for social neuroscience, Ochsner and
Lieberman (2001) argued that social neuroscience would do for
social psychology what cognitive neuroscience had done for
cognitive psychology, "as data about the brain began to be used
to constrain theories about... cognitive processes...."
For example, in an early paper,
Cacioppo and Berntsen (1992) asserted that "knowledge of the
body and brain can usefully constrain and inspire concepts and
theories of psychological function." In
a more recent advertisement for social neuroscience, Ochsner and
Lieberman (2001) argued that social neuroscience would do for
social psychology what cognitive neuroscience had done for
cognitive psychology, "as data about the brain began to be used
to constrain theories about... cognitive processes...."
 And Goleman (2006) drew an
analogy to theorizing at the sociocultural level, arguing that
"the basic assumptions of economics... have been challenged by
the emerging 'neuro-economics'... [whose] findings have shaken
standard thinking in economics."
And Goleman (2006) drew an
analogy to theorizing at the sociocultural level, arguing that
"the basic assumptions of economics... have been challenged by
the emerging 'neuro-economics'... [whose] findings have shaken
standard thinking in economics."
 To some extent, Fiske and
Taylor (2013) appear to have bought into this idea, where they
write that "Brains matter..." because "social-cognitive
processes can be dissociated on the basis of distinct
neuroscientific responses". By which they seem to mean
that neuroscientific evidence can identify different cognitive modules underlying social
cognition.
To some extent, Fiske and
Taylor (2013) appear to have bought into this idea, where they
write that "Brains matter..." because "social-cognitive
processes can be dissociated on the basis of distinct
neuroscientific responses". By which they seem to mean
that neuroscientific evidence can identify different cognitive modules underlying social
cognition.
With
respect to the Goleman quote, it has
to be said that the basic assumptions of economics were shaken,
all right, but not by any data about the brain. They were
shaken, first, by Herbert Simon's observational studies of
organizational decision-making, which led him to propose satisficing
as a way of coping with bounded rationality; and
Kahneman and Tversky's questionnaire studies of various kinds of
choices, which led them to their heuristics and biases
program for studying judgment and decision-making, and prospect
theory as an alternative to traditional rational choice
theory.
 It is instructive,
as well, to consider how the rhetoric of constraint worked out
in cognitive neuroscience -- which, after all, is what inspired
social neuroscience in the first place. Cognitive
neuroscientists often cite cases of hippocampal amnesia (such as
Patient H.M.) as an example of how neuroscientific findings led
to a revolution in theories of memory. While it is true
that studies of H.M. and other amnesic patients, as well as
neuroimaging studies, showed that the hippocampus played an
important role in memory, the explanation of precisely what that
role was has changed greatly over the years. In each case,
the explanation of the function of the hippocampus followed,
rather than preceded, an advance in the theory of memory.
It is instructive,
as well, to consider how the rhetoric of constraint worked out
in cognitive neuroscience -- which, after all, is what inspired
social neuroscience in the first place. Cognitive
neuroscientists often cite cases of hippocampal amnesia (such as
Patient H.M.) as an example of how neuroscientific findings led
to a revolution in theories of memory. While it is true
that studies of H.M. and other amnesic patients, as well as
neuroimaging studies, showed that the hippocampus played an
important role in memory, the explanation of precisely what that
role was has changed greatly over the years. In each case,
the explanation of the function of the hippocampus followed,
rather than preceded, an advance in the theory of memory.
As a general rule, it seems that the proper interpretation of neuroscientific data ultimately depends on the availability of a valid psychology theory of the task being performed by the patient or the subject. The constraints go down, not up -- meaning that psychological theory constrains the interpretation of neuroscientific data. Unless the psychology is right, the neuroscience will be wrong.
Here's one possible counterexample:
Zaki et al. (2011) employed fMRI in an attempt to resole a
longstanding theoretical dispute over the nature of social
influence in Asch's (1956) classic study of conformity. Asch
had found that even in a fairly simple perceptual task, subjects
tended to go along with the incorrect judgments of a unanimous
majority. But Asch himself raised the question of whether
his subjects' behavior reflected private acceptance, or
conformity at the level of perception -- that is, actual changes
in perception in response to social influence -- or merely public
compliance, or conformity at the level of behavior -- that
is, in the absence of any change in the subjects' perceptual
experience. Zaki et al. had their subjects rate the physical
attractiveness of photographs of faces; they rated the faces again
after receiving normative feedback about how their peers also
rated the faces. There was a clear conformity effect:
subjects shifted their ratings in the positive direction if the
peer judgments were higher than their initial judgments, and they
shifted their judgments lower if the peer judgments were
lower. Simultaneous fMRI recordings showed activation in the
nucleus accumbens (NAcc) and orbitofrontal cortex (OFC) during the
second rating session; levels of activation in these areas are
thought to reflect the values assigned to stimuli. were also
activated when the subjects anticipated, and won, monetary
prizes. Zaki et al. concluded that conformity "is
accompanied by alterations in the neural representation of value
associated with stimuli" (p. 898) -- i.e., that conformity in
their experiment was at the level of perception, not merely at the
level of behavior. At the same time, the NAcc and OFC
perform many and varied functions. For example, the NAcc is
involved in the processing of reward and aversion, and might well
be activated just by looking at faces that are rated as especially
attractive or unattractive. And the OFC is important for
decision-making, and might be activated by the process of
considering the relationship between the subjects' judgments about
the faces, and those of their ostensible peers. In any
event, the functions of both NAcc and OFC are both determined by
behavioral and self-report evidence in the first place, so it
can't be said that the neuroscientific facts psychological
theories all on their own.
And another one: Robin Dunbar (2014)'s social-brain hypothesis holds that the size of the cerebral cortex permits, and also constrains, the number of social contacts on can have. He argues that our brain size, big as it is (relative to the rest of our bodies) evolved to allow us to deal with social groups of about 150 people. More than that, we just don't have enough neurons to keep track of everyone. He and his colleagues have reported a brain-imaging study which shows that the volume of the ventromedial prefrontal cortex (VMPFC) predicts both the size of an individual's social networks, and the number of people whose mental states we can keep in mind at once (Lewis et al., NeuroiImage 2011). While it's not clear that this result is anything more than a corollary to the well-known limits on short-term or working memory, the social-brain hypothesis at least relies on biological data to set hypothetical upper limits on both the number of acquaintances and levels of intentionality.
Pardon the Dualism, but...Only when we know what the mind is doing can we begin to understand what the brain is doing. |
| Physiology is a tool
for psychology, not an obligation.
(Anonymous, c. 1980) |
|
Psychology isn't just something to do until the biochemist comes (Neisser, 1967). |
|
Psychology without neuroscience is still the science of mental life, but neuroscience without psychology is just the science of neurons. (Kihlstrom, 2010) |
Even though it's doubtful that neuroscientific evidence can contribute to psychological theory, neuroscientific evidence is the only way we have of discovering the neural substrates of cognitive or social processes. And that's enough.
This page last revised 05/21/2019.

